|
Capítulo 4: Tejidos
Definición Regresar
Una consecuencia de la multicelularidad es que posibilita la división del trabajo. La célula de un ser unicelular debe cumplir con todas las funciones inherentes a la vida. En los seres multicelulares las células pueden dividirse el trabajo, y distintos grupos celulares se especializan en distintas funciones, formando tejidos. Un tejido es un conjunto de células similares, asociadas entre sí, que se han adaptado para realizar funciones específicas. Los tejidos diferentes a su vez se ordenan y forman órganos, pudiendo desarrollar una amplia variedad de tareas.
Cuanto más especializada está una célula en una función determinada, más eficiente se hace para cumplir con ese fin. La especialización conlleva una optimización de la estructura y la función, pero puede ir en detrimento de otras capacidades. Las células que forman parte de una comunidad pluricelular pierden las características necesarias para vivir en forma independiente. Por ejemplo, las neuronas son células con alto grado de especialización y una estructura tan compleja, que, en general, no se dividen, pues al dividirse perderían las adaptaciones logradas.
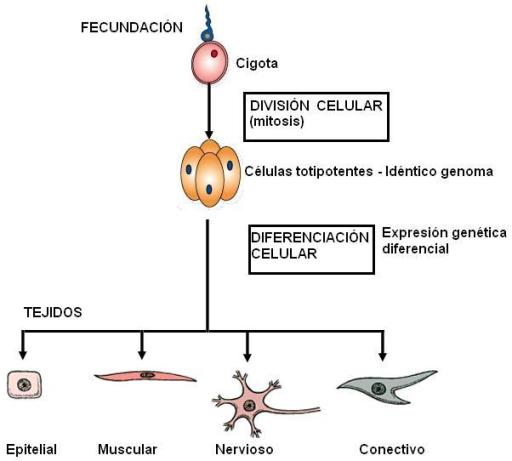
Todas las células que forman el organismo humano provienen de una célula originaria, la cigota, producto de la fusión de un óvulo y un espermatozoide. La cigota experimenta mitosis sucesivas formando un embrión. Como la mitosis genera células hijas con idéntica información genética, todas las células comparten el mismo genoma. En los primeros estadios del embrión, las células son totipotentes. De hecho, si se las separa en dos grupos, cada uno puede generar un organismo completo (este fenómeno es el que da origen a los gemelos monocigóticos). Sin embargo, muy pronto las células embrionarias inician el proceso de diferenciación. Complejos mecanismos hacen que se “enciendan” o “apaguen” determinados conjuntos de genes en diferentes grupos de células y así las células van adquiriendo diferentes características y forman los tejidos. Una vez diferenciadas, las células pierden su totipotencialidad. Significa que conservarán la identidad adquirida para sí y sus descendientes y en general no podrán convertirse a otro tipo celular. Por eso, si se separan las células de un embrión en una etapa avanzada del desarrollo, cada parte ya no está en condiciones de formar la parte que le falta. Del mismo modo, un adulto no puede reemplazar neuronas lesionadas a partir de otro tipo celular.
En el organismo humano se han reconocido más de 200 tipos celulares distintos; sin embargo, los grupos celulares pueden reducirse a cuatro tejidos básicos: epitelial, conectivo, muscular y nervioso.
Las características de un tejido no solo dependen de las células que lo componen. En las propiedades de un tejido tiene un papel muy importante el material que rodea a las células: la sustancia intercelular o matriz extracelular (MEC).
A continuación se analizarán los componentes y funciones de los cuatro tejidos principales.
Tejido epitelial Regresar
El tejido epitelial es el encargado de tapizar las superficies y cavidades corporales. Se lo encuentra en la piel, en las membranas que revisten órganos, como el peritoneo, las pleuras y el pericardio (que recubren a los órganos abdominales, los pulmones y el corazón, respectivamente) y tapizando el interior de los órganos huecos, como el estómago o los vasos sanguíneos.
Una característica sobresaliente del tejido epitelial es su integridad; los epitelios son membranas, capas continuas de células. Esto se debe a que la sustancia intercelular es escasa y las células epiteliales se mantienen estrechamente unidas entre sí por medio de una serie de diferenciaciones de membrana que conforman el “complejo de unión”.
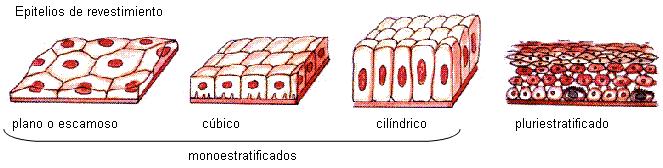
Los epitelios se clasifican según la forma de sus células, en: escamosos, cúbicos y columnares o cilíndricos.
Pueden ser monoestratificados (formados por una capa de células) o pluriestratificados (cuando tienen varias capas celulares).
Los epitelios monoestratificados suelen estar asociados a funciones de intercambio de sustancias; los pluriestratificados, en cambio, están presentes en superficies más expuestas a agresiones físicas, como la piel.
 |
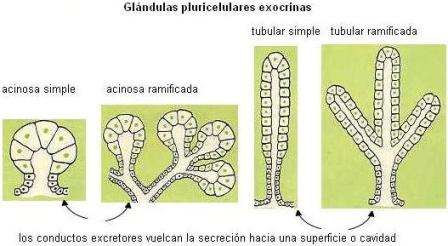
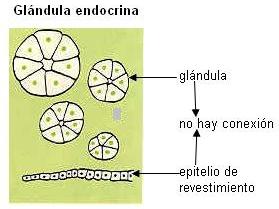
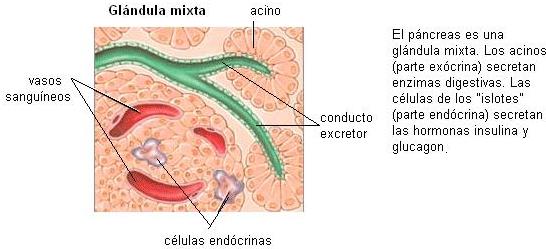
Algunos tipos de células epiteliales se especializan en la secreción de sustancias: son células glandulares. Las células glandulares pueden estar dispersas entre otras células de una membrana epitelial o pueden formar órganos especializados llamados glándulas. Tanto las glándulas unicelulares como las pluricelulares se clasifican en exocrinas y endocrinas.
Una glándula exocrina es la que vuelca su secreción a una cavidad corporal o a la superficie corporal. Las glándulas exocrinas presentan conductos excretores, que las conectan con estas superficies.
Las glándulas endocrinas carecen de conductos excretores; sus secreciones se vuelcan al medio interno y son transportadas por la sangre. Estas secreciones se denominan “hormonas”.
Diferenciaciones de membrana Regresar
Las diferenciaciones de membrana son regiones de la membrana plasmática especializadas en diferentes funciones.
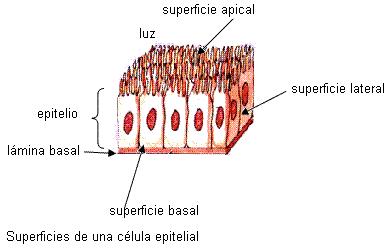
Las células del tejido epitelial ofrecen tres superficies distintas: apical, lateral y basal. La superficie apical está en contacto con la luz, en los órganos huecos, o con alguna cavidad corporal. Mediante la superficie lateral, las células epiteliales contactan entre sí. La superficie basal siempre se apoya sobre un tejido conectivo. La matriz extracelular presenta, inmediatamente por debajo de la superficie basal, una zona denominada “lámina basal”. En cada una de estas superficies de las células epiteliales hay diferenciaciones de membrana adaptadas a funciones distintas.
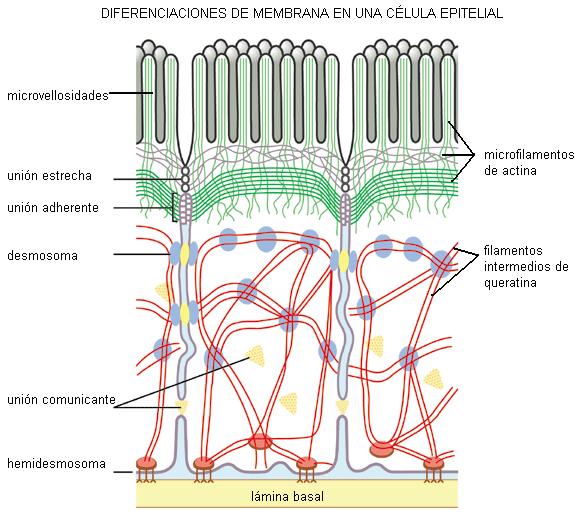 |
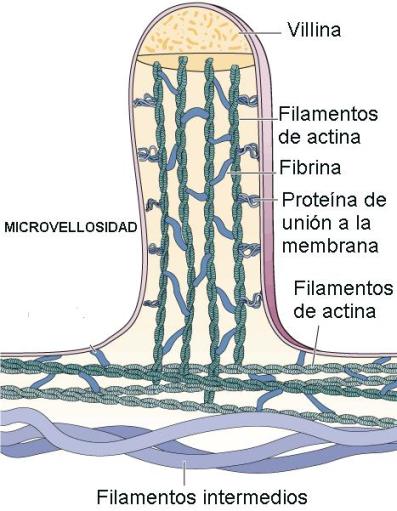
1) Microvellosidades
Son diferenciaciones de la superficie apical, a modo de delgados pliegues. Las microvellosidades poseen en su interior un sostén de microfilamentos en una estructura muy ordenada.
Las microvellosidades no son móviles. Su función consiste en proporcionar un área mayor de membrana en poco espacio, a fin de optimizar la absorción de sustancias. Por ejemplo, en las paredes del intestino delgado, las microvellosidades aumentan la superficie para la absorción de nutrientes.
2) Complejos de unión
Los complejos de unión son un conjunto de diferenciaciones de la superficie lateral de las células epiteliales, que sellan el espacio entre las células y mantienen la integridad del epitelio. El complejo de unión incluye a las uniones estrechas, las uniones adherentes y los desmosomas.
• Unión estrecha (unión oclusiva o zonula occludens)
Las uniones estrechas son regiones de contacto muy íntimo entre las superficies laterales de las células. Al microscopio electrónico se observan como zonas donde las membranas celulares de las células adyacentes se tocan sin dejar espacio intercelular. El contacto se establece por medio de las proteínas de membrana “ocludinas” y “claudinas” que forman cordones selladores o “costuras” entre las células vecinas. Por su cara citosólica estas proteínas contactan con el citoesqueleto de actina.
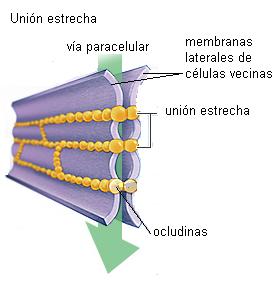 |
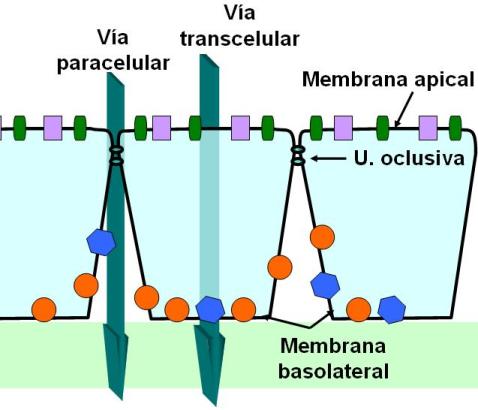
Los epitelios se comportan como barreras entre dos medios. Las uniones estrechas contribuyen a hacer de los epitelios barreras selectivas. Ocluyen el espacio intercelular impidiendo el pasaje de moléculas aun pequeñas por dicho espacio. De esta forma, toda sustancia que atraviese el epitelio deberá hacerlo a través de las células (vía transcelular) y no entre las células (vía paracelular).
Por otro lado, las uniones estrechas bloquean el movimiento de las proteínas que forman la membrana. Impiden que proteínas de membrana de la superficie basolateral se desplacen hacia la superficie apical y viceversa. La consecuencia es que el epitelio se “polariza”, es decir, posee diferentes tipos de proteínas en ambas superficies, permitiendo que cada una cumpla una función distinta. Por ejemplo, en el epitelio intestinal, la superficie apical tiene transportadores de glucosa que introducen este nutriente desde la luz hacia las células, mientras que la membrana basal tiene otro tipo de transportadores que exportan la glucosa hacia los vasos sanguíneos que están en contacto con esta superficie. Así se logra el transporte de la glucosa desde la luz hacia la sangre.
 |
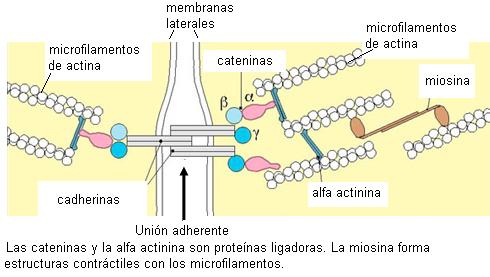
• Unión adherente (zonula adherens)
Las uniones adherentes contribuyen a dar cohesión al tejido. Se encuentran en la superficie lateral formando un cinturón que rodea a la célula y la mantiene adherida a las células vecinas. La adhesión depende de unas proteínas de membrana llamadas “cadherinas”. Éstas presentan una zona o dominio que se proyecta hacia el exterior, haciendo contacto con las cadherinas de la célula adyacente, como si fueran los dientes de un cierre. El dominio de las cadherinas que se proyecta hacia el citosol se conecta con microfilamentos, los cuales cumplen con funciones de fijación y contracción.
 |
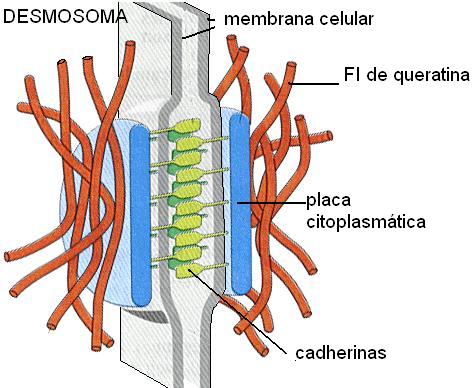
• Desmosoma (macula adherens)
Los desmosomas han sido comparados con “remaches” que unen fuertemente las membranas de células adyacentes. El contacto entre las células está mediado por cadherinas. En la cara citosólica de la membrana, las cadherinas se unen a una placa densa formada por desmina (una proteína ligadora). La desmina establece contacto con filamentos intermedios de queratina. Los desmosomas proporcionan resistencia a las tensiones a que pueden verse sometidos los epitelios.
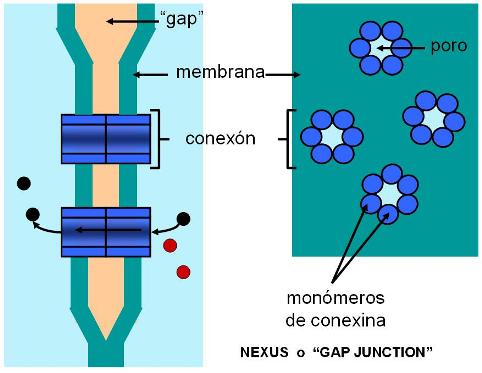
3) Uniones comunicantes (nexus o gap-junction)
Proteínas transmembrana llamadas conexinas determinan un canal o conexón, que al alinearse con una estructura idéntica de la célula vecina forma un canal acuoso que conecta ambos citoplasmas. A través de los conexones las células intercambian iones y pequeñas moléculas. Las uniones de tipo gap son muy importantes en la comunicación intercelular, pues permiten el pasaje de señales eléctricas o químicas que contribuyen al acoplamiento o coordinación entre las células de un tejido.
 |
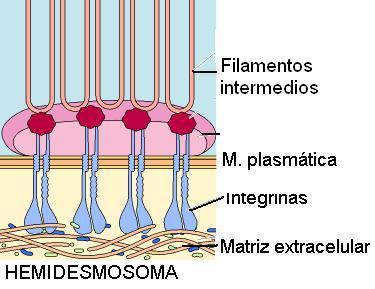
4) Hemidesmosomas
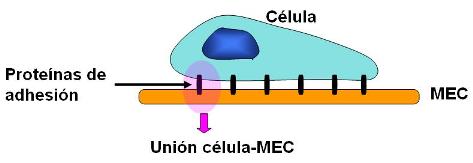
Los hemidesmosomas son uniones de anclaje entre las células y la matriz extracelular (MEC). Se encuentran en la superficie basal de los epitelios, donde éstos establecen contacto con una zona especializada de la MEC, llamada lámina basal.
Su nombre deriva de su semejanza con los desmosomas, aunque solo corresponden a la mitad de estos últimos.
En la unión de la célula con la matriz intervienen proteínas transmembranares de la familia de la “integrinas”. Las integrinas se unen con los microfilamentos del citoesqueleto, por un lado, y con componentes de la MEC por el otro; entre ellos, laminina y colágeno.
Tejido conectivo o conjuntivo Regresar
La característica fundamental del tejido conectivo es el gran desarrollo que en el mismo adquiere la MEC. Los componentes de la MEC son producidos, secretados y orientados por las células del tejido conectivo, llamadas fibroblastos. Los fibroblastos quedan englobados por la MEC que ellos producen.
El tejido conectivo propiamente dicho es el encargado de dar apoyo y nutrición a los epitelios y de conectar entre sí a otros tejidos y órganos.
Además del tejido conectivo propiamente dicho, existen otros tejidos más especializados, que derivan de aquél, como el óseo, el cartilaginoso y el sanguíneo.
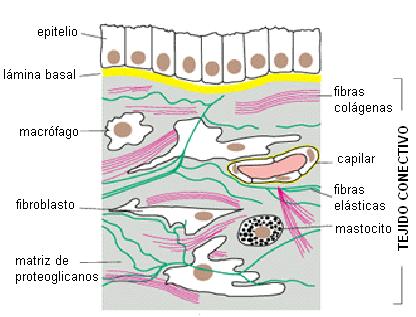 |
La matriz extracelular del tejido conectivo contiene una sustancia fundamental, amorfa, en la cual se ubican componentes fibrosos.
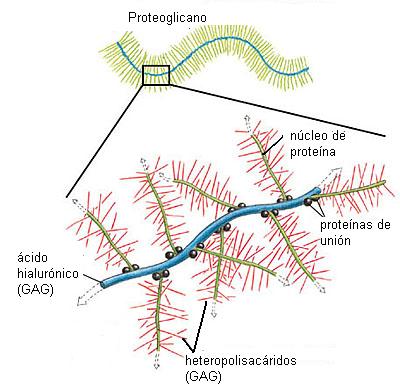
La sustancia fundamental es un gel de proteoglicanos. Los proteoglicanos son macromoléculas formadas por heteropolisacáridos (también llamados GAG: glucosa-amino-glicanos) y proteínas. Se trata de moléculas con abundancia de grupos polares, con cargas negativas, las cuales ejercen gran atracción por el agua y el Na+. Son un excelente medio para la difusión de nutrientes y de otras moléculas que actúan como mensajeros químicos. A su vez, por la gran retención de agua, están capacitados para resistir fuerzas de compresión, comportándose como una esponja.
 |
Embebidos en la matriz de proteoglicanos se encuentran los elementos fibrosos del tejido conectivo: las fibras elásticas y las fibras colágenas.
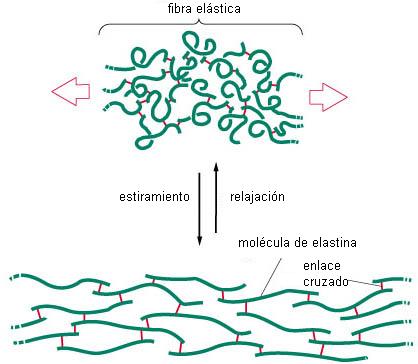
Las fibras elásticas están constituidas por la proteína elastina, que se dispone en forma de láminas y redes, mantenidas por enlaces cruzados. Las fibras elásticas son extensibles y tienen la capacidad de recobrar su conformación inicial después de una deformación transitoria. Son muy abundantes en las paredes arteriales.
 |
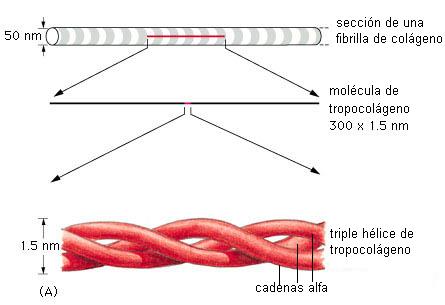
Las fibras colágenas son rígidas y se oponen a las fuerzas de tracción. La unidad molecular que forma las fibras colágenas es el tropocolágeno.
Una molécula de tropocolágeno está formada por tres cadenas de polipéptido, llamadas cadenas alfa. Existen más de 20 tipos de cadenas alfa, cuyas combinaciones dan origen a muchos tipos de colágeno.
Las cadenas alfa tienen una estructura primaria muy peculiar, con una alta proporción de residuos de los aminoácidos glicina y prolina, en secuencias repetitivas. Cada cadena alfa adopta la forma de una hélice de giro izquierdo; las tres juntas se enrollan hacia la derecha, formando una superhélice, de manera similar a las hebras que componen una soga.
Las moléculas de tropocolágeno tienden a alinearse y las hileras a su vez forman haces o fibrillas. Varias fibrillas conforman una fibra. Las fibras de colágeno se disponen en distintos ordenamientos, formando redes o láminas.
 |
En la matriz extracelular están presentes otras proteínas que cumplen función de adhesión, por ejemplo, la laminina y la fibronectina. Estas proteínas interactúan, por un lado, con integrinas de la membrana celular y por otro, con componentes de la matriz. Las proteínas de adhesión participan en las uniones célula-matriz y célula-célula. La fibronectina falta o está reducida en células cancerosas. Esto podría explicar la tendencia de estas células a romper sus conexiones con las células vecinas e invadir tejidos distantes (metástasis).
 |
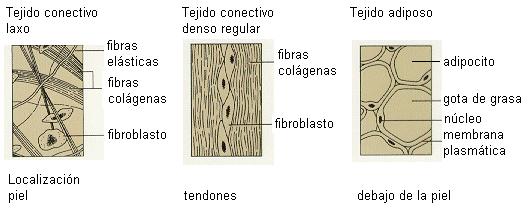
La proporción entre el componente amorfo y el fibroso y el ordenamiento de este último en la MEC da lugar a las distintas variedades del tejido conectivo propiamente dicho.
El tejido conectivo laxo tiene un predominio del componente amorfo, mientras que el denso es más rico en fibras. A su vez, los conectivos densos pueden ser irregulares o regulares. El tejido denso regular posee sus fibras ordenadas en la dirección en que el tejido sufre fuerzas de tracción; se encuentra, por ejemplo, en los tendones.
El tejido adiposo es una variante del tejido conectivo laxo. Las células se diferencian a adipocitos, que están adaptados a la reserva de grasas.
Tejido muscular Regresar
El tejido muscular está formado por escasa sustancia intercelular y células muy diferenciadas, llamadas “fibras”, especializadas en la contracción. El aparato contráctil de las células musculares está constituido por microfilamentos de actina asociados con miosina, en estructuras sumamente ordenadas.
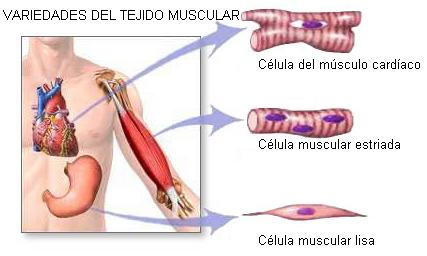
Existen tres variedades de tejido muscular: el estriado, el cardíaco y el liso.
El tejido muscular estriado está formado por células cilíndricas multinucleadas cuyos núcleos tienen una posición periférica. Se ubica en los músculos esqueléticos.
El tejido muscular cardíaco, también estriado, posee células anastomosadas, multinucleadas, con núcleos en posición central. Se localiza en el corazón, formando el miocardio.
El tejido muscular liso consta de células fusiformes y con un solo núcleo. Se lo encuentra en las paredes de arterias y venas y de los órganos huecos, como el tubo digestivo, la vejiga urinaria, etc.
Tejido nervioso Regresar
El tejido nervioso está formado por neuronas, células especializadas en la conducción de impulsos nerviosos, y células gliales o “de la glía”, que dan sostén y nutrición a las neuronas.
Una neurona típica tiene un cuerpo celular, donde se ubica el núcleo, y dos tipos de prolongaciones: las dendritas y el axón.
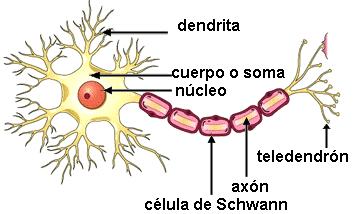 |
| Las células de Schwann pertenecen a la glía. Recubren el axón de ciertas neuronas. |
| Resumen de los tejidos principales |
| Tejido |
Células |
Sustancia intercelular |
Función |
Epitelial |
Poco diferenciadas |
Escasa |
Revestimiento y secreción |
Conectivo |
Poco diferenciadas |
Abundante |
Conexión entre tejidos y órganos |
Muscular |
Muy diferenciadas |
Escasa |
Contracción |
Nervioso |
Muy diferenciadas |
Escasa |
Conducción de impulsos |
|
|