|
Capítulo 3: Célula
Introducción al estudio de la célula
Teoría celular Regresar
La teoría celular se debe a dos científicos alemanes, el botánico Mathias Schleiden y el zoólogo Theodor Schwann. En 1838, Schleiden señaló por primera vez que las plantas se componen de células. Al año siguiente, Schwann extendió esta generalización a los animales. La teoría celular no tardó en imponerse, pues agrupó un conjunto de datos que ya gozaban de consenso en la comunidad científica y desde entonces se acepta que la célula es la unidad básica de todos los organismos vivos.
En el año 1855, Rudolfh Virchow amplió la teoría celular y afirmó que las células solo surgen por división de otras células preexistentes, contradiciendo así la teoría (que aún entonces tenía muchos adeptos), de que las células pueden surgir por generación espontánea de la materia inanimada.
Durante el siglo XX, la teoría celular fue reafirmada y ampliada y es hoy uno de los conceptos unificadores más importantes de la biología. En su formulación actual, la teoría celular enuncia:
1) Los seres vivos están formados por células y productos celulares.
2) Las células se originan a partir de otras células.
3) Las reacciones químicas del organismo vivo tienen lugar dentro de células.
4) Las células contienen la información hereditaria de los organismos que integran y esta información se transmite de la célula madre a la célula hija.
Características de las células Regresar
Todas aquellas características que se hacen evidentes en un organismo complejo y nos permiten reconocerlo como un ser vivo, están presentes en cada una de las células que lo componen.
Las características de las células son:
· Tienen una organización compleja.
· Son sistemas abiertos: intercambian materia y energía con el medio.
· Realizan una serie de transformaciones químicas a las cuales se les da el nombre de metabolismo.
· Poseen un programa genético que guía el desarrollo de sus estructuras y su funcionamiento. Ese programa genético está inscripto en la estructura del ADN (ácido desoxirribonucleico) y contiene información para la síntesis de proteínas. Sin embargo, el ADN no participa en forma directa en la elaboración de proteínas. Para ello, la célula sintetiza una molécula intermediaria, el ARN (ácido ribonucleico), donde se transcribe la información genética almacenada en el ADN. El ARN es el artífice directo de la síntesis de proteínas, proceso también llamado traducción. Las proteínas son las ejecutoras del programa. Por lo tanto, la puesta en marcha de un programa genético requiere:
 |
. Tienen movimiento.
· Poseen receptores que les permiten captar señales del medio y responden a ellas.
· Se autorregulan.
· Se reproducen.
Tamaño celular Regresar
Las células miden típicamente unos pocos micrómetros (=1mm = 10-6m) de diámetro. Las células no sobrevivirían con volúmenes mayores. El límite al tamaño celular viene impuesto fundamentalmente por dos necesidades:
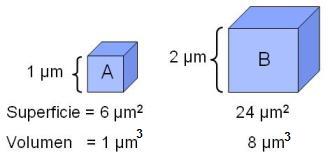
· Que la superficie celular, a través de la cual se realizan los intercambios con el medio, dé abasto para suministrarle a la célula los nutrientes necesarios y permitirle la eliminación de sus desechos. Cuanto mayor es el volumen de un cuerpo, proporcionalmente menor resulta su superficie. Tómese este simple ejemplo: un cubo A de 1 micrómetro de lado tiene una superficie de 6 micrómetros cuadrados y un volumen de 1 micrómetro cúbico. Un cubo B de 2 micrómetros de lado tiene una superficie de 24 micrómetros cuadrados y un volumen de 8 micrómetros cúbicos. Mientras la superficie de B sólo cuadruplica la de A, su volumen es 8 veces mayor. Si la superficie de estos cuerpos tuviera que ser utilizada para realizar intercambios, como ocurre con las células, el cuerpo B sería menos eficiente que A, ya que dispone de una superficie relativamente menor para su volumen.
 |
· Que el volumen celular sea lo suficientemente pequeño para que las moléculas que participan del metabolismo puedan llegar de una parte a otra de la célula en un tiempo breve.
Unidades de longitud utilizadas en Biología celular Regresar
- Tamaño celular: se expresa en micrómetros (antes llamados micrones).
- Tamaño de estructuras subcelulares: se expresa en nanómetros = milimicrones.
- Tamaño de macromoléculas: se expresa en angstroms.
1 micrómetro = 1mm = 10-6 m
1 nanómetro = 1nm = 10-9 m
1 angstrom = 1 Aº = 10-10 m |
Microscopios Regresar
El poder de resolución de un instrumento óptico es la capacidad de discriminar o “ver” separadamente dos puntos. El límite de resolución es la menor distancia que debe separar a dos puntos para que el instrumento los discrimine como puntos separados. Por lo tanto, la capacidad resolutiva es tanto mayor, cuanto menor sea el límite. El límite de resolución para el ojo humano es 200 mm (=0,2 mm). Salvo algunas excepciones, las células no alcanzan este tamaño, por lo que el estudio de las células requiere la asistencia del microscopio.
Existen dos tipos básicos de microscopio: el microscopio óptico (MO) y el microscopio electrónico (ME). Dados sus pequeños límites de resolución, el poder resolutivo de estos instrumentos (especialmente el del ME) es muy alto y ha permitido el conocimiento detallado de la célula y sus estructuras.
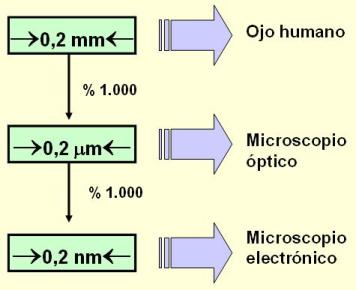 |
El MO posibilita la visión de tejidos y células, con muy poco detalle de su estructura interna.
Las estructuras subcelulares se observan a través del ME. Las imágenes fotográficas además pueden ser ampliadas, lográndose aumentos de hasta10.000.000 x. Al ME es posible observar hasta la estructura general de algunas macromoléculas aisladas, como el ADN. Sin embargo, las estructuras moleculares y los átomos, tal como los hemos dibujado hasta ahora, no pueden ser resueltos por ningún instrumento. Los esquemas que se utilizan son representaciones de modelos teóricos.
Modelos celulares Regresar
Una célula consta de tres elementos fundamentales: un límite, la membrana celular; un contenido o citoplasma y el material genético, el cual se halla en una o más estructuras llamadas cromosomas. El modelo celular más sencillo, el de las bacterias, presenta el cromosoma en contacto directo con el citoplasma, en una zona denominada nucleoide. En todos los demás seres vivos, los cromosomas están encerrados en un núcleo limitado por una envoltura nuclear, de manera que el contenido celular queda dividido en dos zonas: núcleo y citoplasma. A este modelo celular se le dio el nombre de célula eucariota (de eu: verdadero y cario: núcleo). Al tipo celular de las bacterias, en cambio, se lo llamó procariota (anterior al núcleo).
A continuación describiremos la estructura general de una célula eucariota animal típica o idealizada.
Estructura de una célula animal Regresar
Esquema de una célula animal Regresar
Membrana plasmática Regresar
La membrana plasmática o celular es el límite exterior de la célula. Al ME electrónico ofrece una imagen, común con la de otras membranas, llamada “unidad de membrana”.
 |
La descripción de la estructura que sigue no corresponde a una imagen sino a un modelo obtenido de pruebas indirectas.
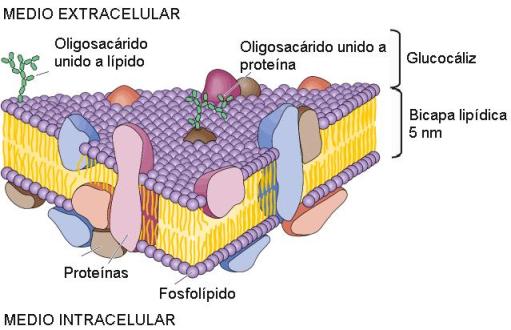
La membrana plasmática es una bicapa formada por fosfolípidos, glucolípidos y colesterol, con proteínas en la superficie de la bicapa e intercaladas entre los lípidos. La mayor parte de las proteínas son en realidad glucoproteínas, pues llevan unidas cadenas de oligosacáridos que se proyectan desde la membrana hacia el exterior celular. Los glucolípidos solo se ubican en la monocapa extracelular y sus glúcidos, por lo tanto, se exponen en la superficie de la célula. Las cadenas glucídicas de unas y otras moléculas forman en conjunto la capa más externa de la membrana plasmática, a la que se llama glucocáliz.
El modelo descriptivo de la membrana plasmática, modelo de mosaico fluido, indica que los componentes de la membrana gozan de cierta libertad de movimiento en el espesor de la misma.
Dependiendo del tejido, la membrana presenta zonas especializadas en distintas funciones, las diferenciaciones de membrana.
 |
La función de la membrana plasmática es, fundamentalmente, el mantenimiento del medio interno de la célula, por medio del control de los ingresos y egresos de moléculas que se producen a través de ella. Algunos de estos pasajes son pasivos, pero otros requieren la intervención de mecanismos de transporte que implican un gasto energético celular. También en la membrana se encuentran estructuras receptoras que le permiten a la célula responder de diversas formas a las señales recibidas. Además, la membrana vincula a una célula con sus vecinas y con la sustancia que la rodea, llamada matriz extracelular. Estas relaciones célula-célula y célula-matriz son de suma importancia en la arquitectura de los tejidos.
Citoplasma Regresar
El citoplasma es la zona que se ubica entre la membrana plasmática y el núcleo. La parte líquida del citoplasma se denomina citosol o matriz citoplasmática. La matriz citoplasmática es un medio acuoso que contiene iones y moléculas pequeñas disueltas y también muchos tipos de macromoléculas en suspensión; entre ellas, enzimas, que participan en importantes vías metabólicas. En algunos casos, el citosol tiene inclusiones, que son depósitos de distintas sustancias. Por ejemplo, en las células del tejido adiposo grandes gotas de triglicéridos se reservan en el citosol.
Sin embargo, el citoplasma está lejos de ser una masa homogénea e informe. Por el contrario, el citosol está interrumpido y recorrido por una serie de estructuras complejas y especializadas: el citoesqueleto, el sistema de endomembranas y los organoides citoplasmáticos.
Citoesqueleto Regresar
El citoesqueleto es el armazón de la célula. Está constituido por tres tipos de elementos filamentosos: los microfilamentos, los microtúbulos y los filamentos intermedios.
Los microfilamentos (MF) son los componentes más delgados del citoesqueleto. Son varillas macizas de 8 nm de diámetro, constituidas por unidades de una proteína globular, la actina G. Las unidades de actina G se unen entre sí en una doble hélice estrecha que forma el microfilamento o actina F.
Los microtúbulos (MT) son cilindros huecos de 25 nm de diámetro; tienen subunidades de otra proteína globular, la tubulina. En la pared del microtúbulo, las unidades de tubulina se disponen formando 13 hileras llamadas protofilamentos.
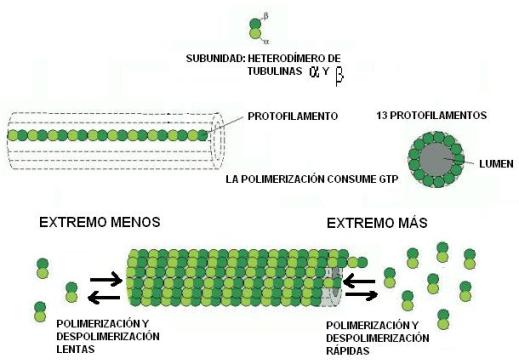 |
Tanto los MF como los MT pueden modificar su longitud añadiendo o perdiendo subunidades por ambos extremos, en los procesos llamados polimerización y despolimerización, respectivamente. La polimerización consume energía.
Microfilamentos y microtúbulos presentan dos extremos que se diferencian en la velocidad de polimerización y despolimerización: en el extremo más estos procesos son rápidos, y en el extremo menos, más lentos. Se dice, por este motivo, que son estructuras polarizadas.
Los microfilamentos se ubican preferencialmente en la zona periférica del citoplasma y pueden adoptar diferentes disposiciones en las células: forman haces, redes delgadas y también redes complejas, tridimensionales. Cambios en los filamentos de actina provocan que la consistencia del citosol pase de sol a gel. La forma en que se disponen los filamentos de actina depende en gran medida de su interacción con las proteínas que se enlazan a actina o proteínas ligadoras. Otras proteínas reguladoras controlan dichos cambios.
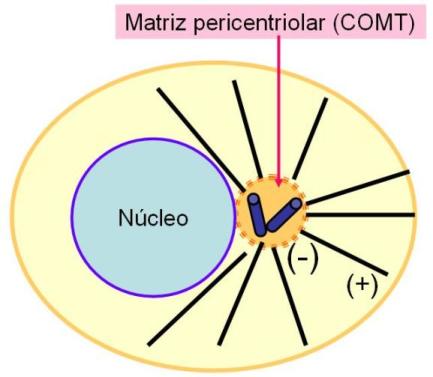
Los MT citoplasmáticos de las células animales generalmente se disponen en el citoplasma en forma de rayos que irradian desde el centrosoma o centro celular. Los extremos menos de los MT se orientan hacia el centrosoma, mientras que los extremos más están dirigidos hacia la periferia celular.
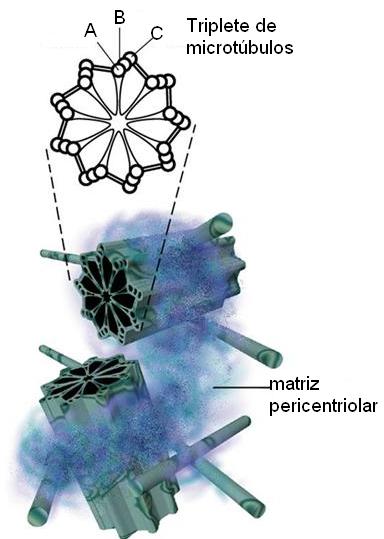
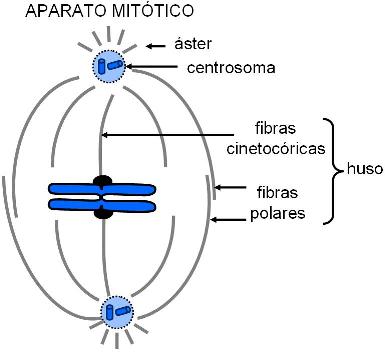
El centrosoma es una zona cercana al núcleo que comprende a los centríolos, estructuras pares ubicadas en ángulo recto uno respecto del otro, y a una matriz que los rodea, la matriz pericentriolar. Esta última contiene proteínas que dirigen la formación y el crecimiento de los microtúbulos, por lo cual el centrosoma es considerado un “centro organizador de microtúbulos” (COMT).
 |
Los MT citoplasmáticos también son estructuras muy dinámicas, con la posibilidad de modificar su longitud y su distribución dentro de la célula. Por ejemplo, durante la división celular, los microtúbulos citoplasmáticos se desensamblan y reorganizan por completo, formando el huso mitótico, estructura encargada de repartir los cromosomas entre las células hijas.
Los microfilamentos interaccionan con proteínas motoras denominadas miosinas I y miosinas II. En asociación con miosinas, los MF participan en el movimiento de organoides, en la migración celular y forman estructuras contráctiles, como el sarcómero de las células musculares.
Los microtúbulos también se asocian a sus propias proteínas motoras, las quinesinas (o cinesinas) y las dineínas. Las quinesinas transportan cargas hacia el extremo más del microtúbulo, y las dineínas transportan sus cargas hacia el extremo menos.
Además de los MT citoplasmáticos, inestables y cambiantes, las células poseen microtúbulos que forman parte de estructuras estables, como los centríolos, las cilias y los flagelos.
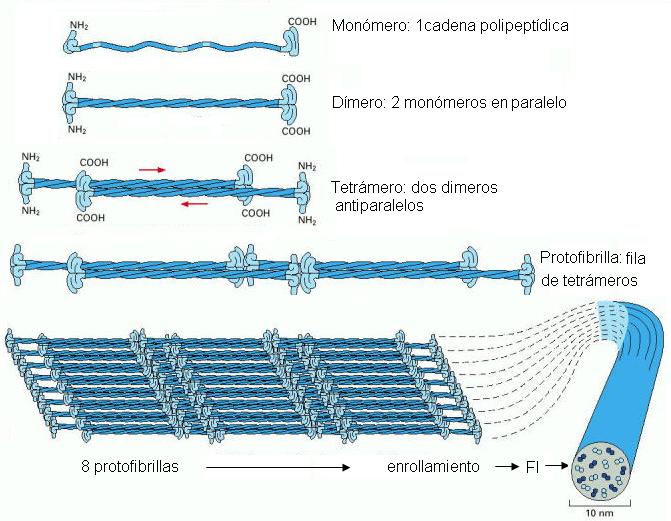
Los filamentos intermedios (FI) tienen un diámetro de 10 nm, intermedio entre el de los microfilamentos y el de los microtúbulos. Se diferencian de MF y MT en que están constituidos por unidades fibrilares y no globulares, y su polimerización ocurre espontáneamente, sin gasto de energía. Tampoco presentan polaridad.
Aunque existen diversas familias de proteínas, específicas de distintos tipos celulares, que conforman los FI, todas ellas están organizadas siguiendo un patrón común.
 |
La función de los FI es netamente estructural. Adoptan una disposición transcelular o transversal al eje mayor de la célula, orientándose en la dirección de las fuerzas que actúan sobre ella.
En conclusión, el citoesqueleto es una estructura dinámica, que cumple con las siguientes funciones: da forma y sostén a la célula, le confiere resistencia a la tracción, participa en el movimiento de organelas dentro del citoplasma y es el responsable del desplazamiento celular.
Sistema de endomembranas Regresar
El sistema de endomembranas (SE) es un enorme compartimiento dentro del citoplasma, como un sistema de cañerías que se interconectan y cuyas paredes están formadas por membrana.
El sistema de endomembranas se encarga de la síntesis de diversas macromoléculas que luego son transportadas por su interior.
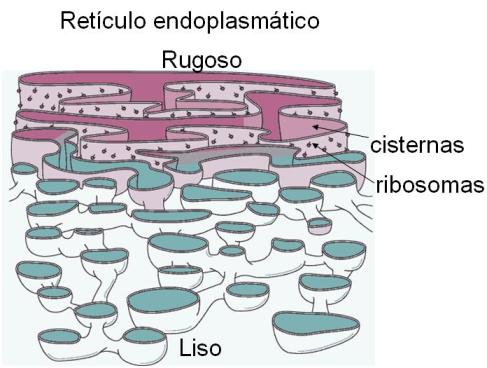
El retículo endoplasmático rugoso o granular (RER o REG) está formado por cisternas o bolsas aplanadas, conectadas unas a otras mediante túbulos membranosos. Sobre la cara de las membranas que da al citoplasma se apoyan ribosomas. Las proteínas sintetizadas por estos orgánulos atraviesan las membranas del REG. Algunas de ellas son volcadas hacia el lumen o cavidad del retículo, mientras que otras quedan ancladas a la membrana. Para que los ribosomas se unan al retículo, la proteína que están sintetizando debe llevar un tipo de señal; de lo contrario, el ribosoma permanece en el citosol, como ribosoma libre. Las proteínas que llevan la señal adecuada y consiguientemente son sintetizadas en ribosomas que se adhieren al retículo, son las siguientes: proteínas integrantes del sistema de endomembranas, proteínas lisosomales, proteínas destinadas a la membrana plasmática y proteínas de secreción.
Al mismo tiempo que las cadenas peptídicas son translocadas hacia el lumen del REG sufren casi siempre un proceso de glicosilación, que consiste en la adición de una o más moléculas de oligosacárido.
El retículo endoplasmático liso o agranular (REL o REA) está representado por túbulos intrincados. En las paredes de estos túbulos se fabrican distintos tipos de lípidos, por ejemplo fosfolípidos de membrana y esteroides. La cavidad o luz del REL también sirve como reservorio de calcio iónico. Otra función del REL, particularmente en las células hepáticas, es su acción detoxificante.
 |
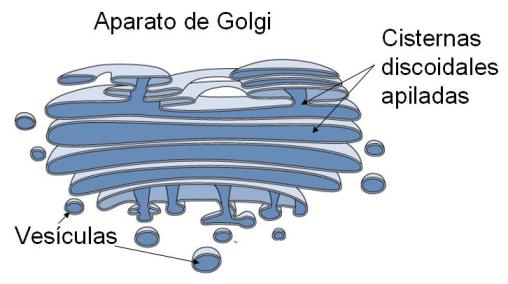
El aparato o complejo de Golgi consta de bolsas circulares apiladas unas sobre otras. Allí se sintetizan algunos glúcidos complejos (heteropolisacáridos) y además se reciben vesículas de transporte con las moléculas elaboradas por el REG y el REL. Las vesículas o vacuolas son pequeñas bolsas membranosas que se desprenden como brotes de los dos sectores del retículo endoplasmático y sirven como “empaque” y vehículo de transporte para los productos allí fabricados. Las vesículas de transporte son desplazadas por el citosol hasta llegar al aparato de Golgi. Este complejo presenta una cara de recepción (cara cis), a la cual se fusionan las vesículas, entregando su cargamento. Ya dentro del aparato de Golgi, los distintos cargamentos son modificados, concentrados y clasificados. Así, según su destino, son empacados en nuevas vesículas que brotan de la cara opuesta del aparato de Golgi, la cara de emisión (cara trans). Algunas de las vesículas emitidas por el aparato de Golgi son lisosomas; otras son vesículas de secreción.
 |
Los lisosomas contienen las proteínas lisosomales originadas en el REG. Las mismas son enzimas hidrolíticas y participan en procesos de digestión intracelular.
Las vesículas de secreción se fusionan con la membrana plasmática. Esto permite volcar su contenido en el medio extracelular, donde cumplirá una función, ya sea hormonal, enzimática, estructural, etc.
La fusión de vesículas con la membrana plasmática también es el medio para que ésta incorpore nuevos componentes, lípidos o proteínas, sintetizados en el sistema de endomembranas y transportados como parte integrante de la membrana vesicular.
Ribosomas Regresar
Estos orgánulos están formados por dos subunidades, la mayor y la menor, que se ensamblan entre sí en presencia de un tipo de ARN llamado mensajero (ARNm). Cada ARNm es una molécula lineal que porta la información para la síntesis de una proteína particular. El ribosoma ya ensamblado se desliza sobre el ARNm, que se sitúa en un túnel excavado en la subunidad menor, al tiempo que sintetiza la proteína especificada en el ARN. Según las señales que exhiben las proteínas nacientes, el ribosoma persiste en el citosol, como ribosoma libre, o se adhiere a las membranas del REG y continúa allí el proceso de síntesis, como ya se mencionó. Las subunidades ribosomales se separan una vez que la síntesis de la proteína ha concluido.
Frecuentemente, varios ribosomas, ya sean libres o unidos al REG, aparecen agrupados sobre un mismo ARNm; a estos grupos se los denomina polisomas o polirribosomas.
Los ribosomas carecen de membrana y cada una de sus subunidades es un complejo formado por ARN ribosomal (ARNr) y proteínas.
Mitocondrias Regresar
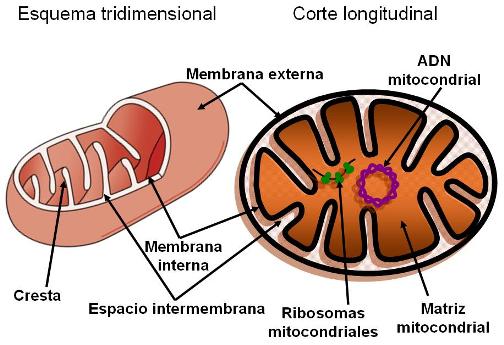
Las mitocondrias tienen forma ovoide, de bastón o esférica. Son organoides de gran tamaño (hasta 3 micrómetros de largo). Están rodeadas por dos membranas: una externa, lisa y otra interna, con pliegues llamados crestas mitocondriales. Entre ambas membranas hay un compartimiento, el espacio intermembrana, y por dentro de la membrana interna se encuentra la matriz mitocondrial. Sobre la membrana interna y en la matriz se ubican una gran cantidad de enzimas que participan en la fase dependiente de oxígeno de la respiración celular. La función de las mitocondrias es, por lo tanto, la de proveer energía para el funcionamiento celular. La energía que se libera de los combustibles durante la respiración celular se almacena temporariamente en una molécula llamada ATP (sigla del nucleótido adenosín trifosfato). La mitocondria es la principal proveedora de ATP de la célula.
Las mitocondrias poseen su propio ADN, así como ribosomas (más pequeños que los citoplasmáticos) y otros tipos de ARN. Parte de la información genética necesaria para el funcionamiento de las mitocondrias está contenida en su ADN; además pueden llevar cabo la traducción. Las mitocondrias son autorreplicantes, se originan por división de las preexistentes.
 |
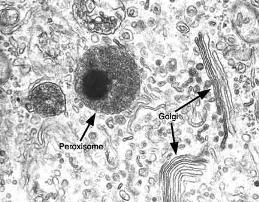
Peroxisomas Regresar
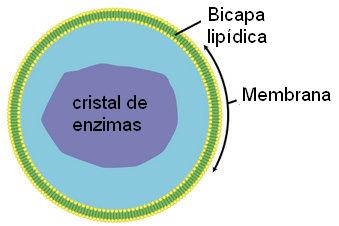
Los peroxisomas son orgánulos membranosos. Contienen enzimas que participan en procesos oxidativos; algunas de ellas tienden a formar un cuerpo cristalino. Son, junto con las mitocondrias, los organoides donde se consume el oxígeno que llega a las células, aunque, a diferencia de lo que ocurre en las mitocondrias, las reacciones que transcurren en los peroxisomas no generan ATP.
Especialmente en células hepáticas, las reacciones oxidativas de los peroxisomas posibilitan la metabolización de sustancias tóxicas, como el etanol o alcohol etílico. El peróxido de hidrógeno (agua oxigenada) se forma como producto de estas reacciones de detoxificación. Una enzima característica del peroxisoma es la catalasa, que cataliza la descomposición del exceso de peróxido de hidrógeno allí generado.
 |
 |
Los peroxisomas no están relacionados con el sistema de endomembranas; cada peroxisoma se origina por división o fisión binaria a partir de otro preexistente y luego crece por incorporación de proteínas captadas selectivamente desde el citosol y lípidos de membrana intercambiados con el REL.
Centríolos Regresar
Los centríolos se encuentran por pares y forman el centrosoma o centro celular junto con la matriz que los rodea (matriz pericentriolar). Desde allí irradian los microtúbulos citoplasmáticos. Los mismos centríolos son cilindros huecos cuyas paredes están formadas por nueve tripletes de microtúbulos. Se duplican antes de la división celular y en el transcurso de ésta se incorporan al aparato mitótico. Al final de la división celular cada célula hija recibe un par de centríolos.
Los centríolos dan origen a las cilias y los flagelos, prolongaciones móviles que están presentes en algunos tipos celulares.
Núcleo celular Regresar
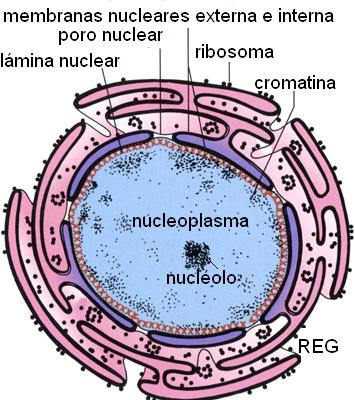
El núcleo está rodeado por una membrana doble, la envoltura nuclear, que es una dependencia del REG; la luz de este último tiene continuidad con la cisterna perinuclear, comprendida entre las dos membranas que forman la envoltura. La envoltura nuclear está sostenida por una estructura de filamentos intermedios, la lámina nuclear.
La envoltura nuclear tiene poros parcialmente cerrados por un complejo proteico, el complejo del poro nuclear (CPN). A través de estos complejos se ejerce el control de las sustancias que ingresan al núcleo o que salen hacia el citoplasma.
Dentro del núcleo se encuentra el jugo nuclear (también llamado nucleoplasma o carioplasma), que es la parte líquida del núcleo, equivalente al citosol. Una matriz o “scaffold” formada por proteínas recorre el nucleoplasma y da sostén a la cromatina.
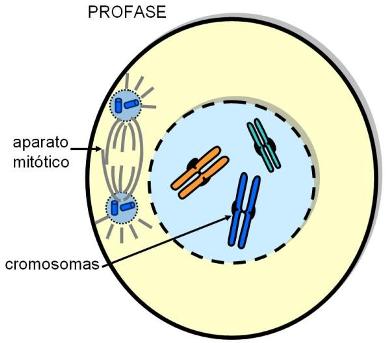
La cromatina es un material fibroso formado por la asociación entre ADN y proteínas, de las cuales las proteínas básicas llamadas histonas son las más abundantes. Las largas moléculas de ADN pueden ser empacadas dentro del núcleo gracias a que se enrollan alrededor de complejos de histonas. Las fibras de cromatina presentan diferentes grados de plegamiento, que resultan en una estructura cada vez más compacta. El grado mayor de plegamiento se alcanza durante la división celular. Entonces la cromatina da lugar a la formación de unas estructuras bien visibles al microscopio óptico, los cromosomas.
El número y el tipo de cromosomas son constantes para cada especie; las células humanas tienen 46 cromosomas.
El nucléolo está formado por la convergencia de varias asas de cromatina en un sector del nucleoplasma. Estas asas son transcritas en moléculas de ARN, precursor del ARNr. A su vez, se acumulan en el nucléolo proteínas provenientes del citosol. Éstas se ensamblan con moléculas de ARNr y así se forman subunidades ribosomales. El nucléolo, entonces, es el sector del núcleo donde se sintetiza ARNr y se arman las subunidades ribosomales. Éstas, una vez construidas, se dirigen al citosol.
 |
l núcleo es la estructura más destacada dentro de la célula, tanto por su tamaño, como por su función. Dentro del núcleo se encuentra el ADN. Pero el núcleo no es solamente el lugar donde se almacenan las instrucciones genéticas; también es el sitio donde se lleva a cabo el primer paso de la expresión genética, que es la transcripción, y donde se ponen en marcha los complicados mecanismos que la regulan. Además, en el interior del núcleo tiene lugar la replicación del ADN, proceso previo y necesario para la división celular.
El núcleo es indispensable para el mantenimiento y la reproducción celular. Si una célula pierde el núcleo muere rápidamente. Aun cuando la pérdida del núcleo está programada como parte del proceso de maduración, como ocurre con los glóbulos rojos, la célula sobrevive muy poco tiempo y es obvio que nunca llega a reproducirse.
Cilias y flagelos Regresar
Las cilias y los flagelos son prolongaciones móviles del citoplasma recubiertas por la membrana plasmática, que se presentan en algunos tipos celulares.
Las cilias son apéndices cortos y numerosos; los flagelos son prolongaciones de mayor longitud y se presentan uno o unos pocos por célula. En el organismo humano, las únicas células flageladas son los espermatozoides. El flagelo forma la cola que propulsa al espermatozoide en el semen y dentro del tracto genital femenino.
Las cilias también actúan como estructuras locomotoras en organismos unicelulares o pluricelulares muy simples, como ciertos protozoos o algas, a los cuales dotan de una locomoción similar al movimiento a remo. Sin embargo, cuando las células ciliadas forman parte de un tejido, las cilias son utilizadas para generar corrientes en el medio que las rodea. El hombre presenta células ciliadas en el epitelio de la vía respiratoria; allí, el movimiento ciliar produce el desplazamiento hacia el exterior de la capa de moco donde quedan atrapadas las partículas que ingresan con el aire. En las trompas de Falopio el movimiento ciliar sirve al fin de atraer el óvulo liberado desde el ovario hacia la cavidad abdominal.
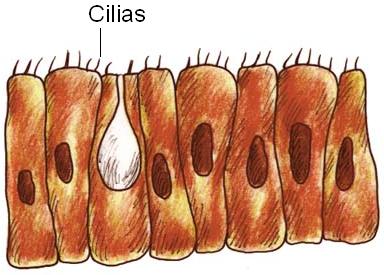 |
Cilias en el epitelio de la vía respiratoria |
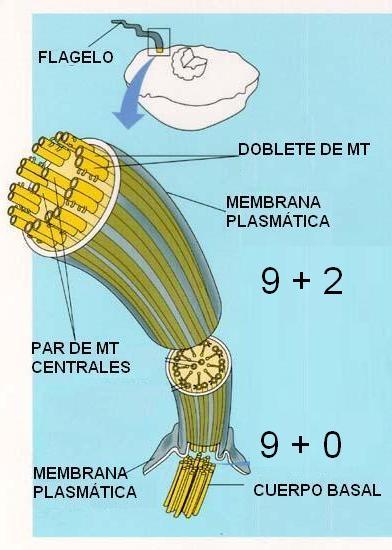
La estructura de las cilias y los flagelos, sin embargo, es la misma. Ambos están rodeados por una prolongación de la membrana plasmática que toma el nombre de axolema. Por dentro se encuentra el axonema, formado por microtúbulos, los que se disponen en nueve dobletes periféricos más un par de microtúbulos centrales (estructura 9+2). Otras proteínas asociadas a los microtúbulos les ayudan a mantener esta organización característica. De cada doblete periférico, además, se proyectan a intervalos regulares dos brazos formados por la dineína ciliar, proteína emparentada con la dineína citoplasmática ya mencionada, que actúa como proteína motora.
En la base de la cilia o el flagelo se ubica el cuerpo basal, formado por nueve tripletes de túbulos. Su estructura se conoce como 9+0 (pues carece de los microtúbulos centrales que se encuentran en el axonema). La estructura del cuerpo basal es igual a la del centríolo. Centríolos y cuerpos basales pueden darse origen unos a otros. Las cilias y flagelos crecen a partir del cuerpo basal.
 |
Naturaleza y flujo de la información genética Regresar
Nucleótidos Regresar
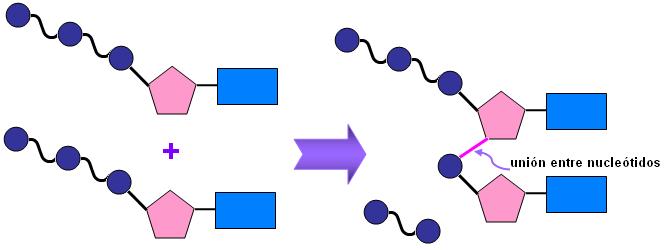
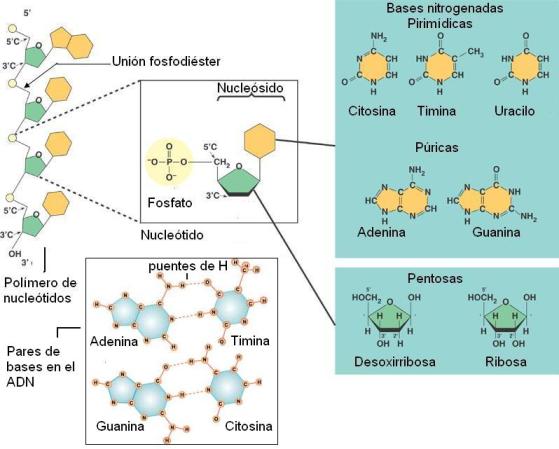
Un nucleótido se forma por la combinación de tres moléculas: una base nitrogenada, una pentosa y ácido fosfórico.
Los nucleótidos pertenecen a dos grupos distintos, según el tipo de pentosa que contienen: los que contienen ribosa son los ribonucleótidos y los que contienen desoxirribosa son los desoxirribonucleótidos.
A su vez, existen dos grupos de bases nitrogenadas: las púricas, formadas por dos anillos de carbono y nitrógeno y las pirimídicas, de menor tamaño que las anteriores, pues constan de un solo anillo. Las bases púricas son adenina (A) y guanina (G) y las pirimídicas incluyen citosina (C), timina (T) y uracilo (U).
La unión de la base nitrogenada con la pentosa forma un nucleósido. A éste pueden adicionársele 1, 2 ó 3 moléculas de ácido fosfórico. Así, se obtienen nucleótidos que se nombran como nucleósidos monofosfato, difosfato o trifosfato (fosfato es el radical que deriva del ácido fosfórico, una vez que éste se une al nucleósido).
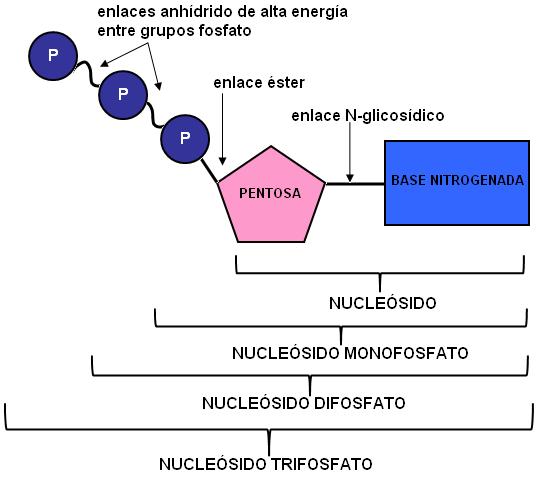 |
Todas las uniones involucradas se producen por reacciones de condensación. La pentosa se enlaza por su C1 a un átomo de nitrógeno de la base nitrogenada, formando un enlace N-glicosídico y por su C5 a un fosfato, mediante un enlace éster.
| NOMENCLATURA DE NUCLEÓSIDOS Y NUCLEÓTIDOS |
| Pentosa |
Base nitrogenada |
Nucleósido |
Nucleótido |
| Monofosfato |
Difosfato |
Trifosfato |
Ribosa
|
Adenina |
Adenosina |
AMP |
AMP |
ATP |
| Guanina |
Guanosina |
GMP |
GMP |
GTP |
| Citosina |
Citidina |
CMP |
CDP |
CTP |
| Timina |
Timidina |
TMP |
TDP |
TTP |
| Uracilo |
Uridina |
UMP |
UDP |
UTP |
Desoxirribosa
(d-Ribosa)
d=desoxi |
Adenina |
d-Adenosina |
d-AMP |
d-ADP |
d-ATP |
| Guanina |
d-Guanosina |
d-GMP |
d-GDP |
d-GTP |
| Citosina |
d-Citidina |
d-CMP |
d-CDP |
d-CTP |
| Timina |
d-Timidina |
d-TMP |
d-TDP |
d-TTP |
| Uracilo |
d-Uridina |
d-UMP |
d-UDP |
d-UTP |
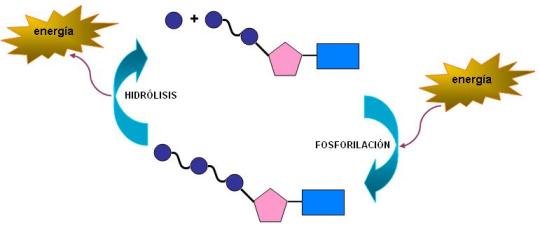
Los fosfatos se unen entre sí por enlaces anhídrido. Los enlaces de tipo anhídrido entre los grupos fosfato pertenecen a un tipo de unión de alta energía. Se los denomina así puesto que almacenan más energía que otros enlaces covalentes. Esto se debe a que los grupos fosfato tienden a ceder protones, adquiriendo cargas negativas que se repelen entre sí; por lo tanto, para unirlos es necesario vencer la fuerza de repulsión, es decir, se debe entregar una cantidad mayor de energía. Inversamente, cuando estas uniones se hidrolizan, la energía es liberada.
A causa de la presencia de los enlaces de alta energía, los nucleósidos difosfato y trifosfato son utilizados como intermediarios energéticos: guardan o ceden una cierta cantidad de energía mediante la formación o hidrólisis de un solo enlace.
 |
Además de la función de intermediarios energéticos, desempeñada principalmente por los ribonucleótidos de adenina (difosfato de adenosina = ADP y trifosfato de adenosina = ATP), los nucleótidos libres son componentes de coenzimas; como tales se unen a ciertas enzimas para cuyo funcionamiento resultan indispensables.
Pero la función de los nucleótidos que nos ocupará en este capítulo es la que cumplen como monómeros de los ácidos nucleicos. Los ribonucleótidos se polimerizan entre sí para formar ácido ribonucleico (ARN), mientras que los desoxirribonucleótidos se polimerizan dando origen al ácido desoxirribonucleico (ADN).
La reacción de polimerización requiere la presencia de nucleótidos trifosfato como sustratos. Sin embargo, al formarse el enlace entre dos nucleótidos, el que se incorpora a la cadena pierde sus dos fosfatos más externos. Así, una vez formados los polímeros, éstos consisten en cadenas de nucleótidos monofosfato.
Ácido ribonucleico (ARN) Regresar
El ácido ribonucleico, o ARN, es un polímero lineal de ribonucleótidos. Los monómeros del ARN son nucleótidos de cuatro clases: de adenina, de guanina, de citosina y de uracilo; en el ARN no hay nucleótidos de timina. Estas cuatro clases de nucleótidos se repiten, en diferentes secuencias, a lo largo de las cadenas de ARN.
Los nucleótidos se enlazan entre sí mediante enlaces fosfodiéster. Dichos enlaces se producen al reaccionar el hidroxilo unido al carbono en posición 3 (C 3’) de la ribosa del primer nucleótido, con el grupo fosfato unido al carbono en posición 5 (C 5’) del nucleótido entrante. Por lo tanto, los dos extremos de la cadena son distintos: el primer nucleótido de la cadena tiene grupo fosfato libre en posición 5’, mientras que el último tiene un hidroxilo libre en posición 3’. Los extremos de la cadena de ARN se nombran entonces como 5’ y 3’, respectivamente.
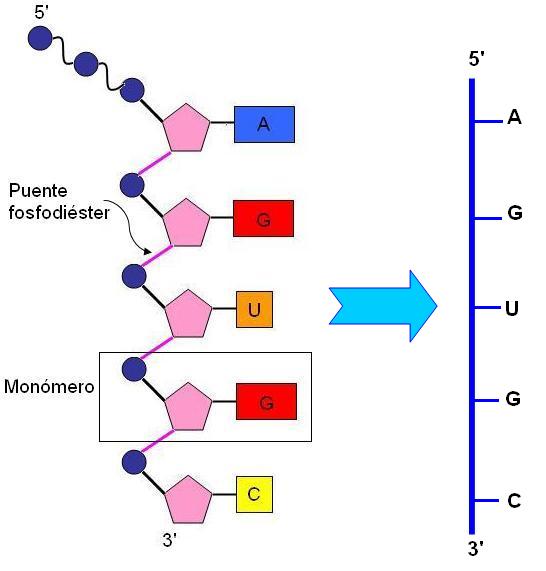 |
Todas las moléculas de ARN comparten esta estructura; sin embargo, hay variaciones que dependen de la longitud de cadena, la secuencia de bases, las modificaciones químicas operadas posteriormente sobre la molécula y el plegamiento o estructura tridimensional que adopta la cadena lineal.
Todos los ARN se sintetizan en el núcleo, pues su síntesis requiere ADN como molde, aunque muchos de ellos son transportados luego al citoplasma.
Existen tres tipos principales de ARN que participan en forma directa en la síntesis de proteínas: el ARN mensajero (ARNm), el ARN ribosomal (ARNr) y el ARN de transferencia (ARNt). Éstos se localizan en el citoplasma.
Otros ARN, llamados ARN pequeños, ubicados tanto en el núcleo como en el citoplasma, participan de diversas formas en procesos relacionados con la maduración y la expresión de los anteriores.
Ácido desoxirribonucleico (ADN) Regresar
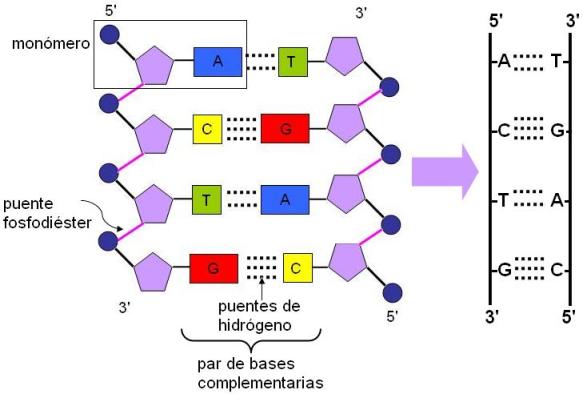
El ADN está formado por dos cadenas de desoxirribonucleótidos. En cada cadena están presentes cuatro clases de nucleótidos: de adenina, de guanina, de citosina y de timina; en el ADN no hay nucleótidos de uracilo. Los nucleótidos de una misma cadena se unen mediante puentes fosfodiéster.
Las dos cadenas que forman la molécula de ADN son antiparalelas, pues están orientadas o “corren” en sentido inverso (5’->3’ y 3’->5’).
Además, ambas cadenas se enfrentan por sus bases, que establecen entre sí uniones del tipo puente de hidrógeno. Los pares de bases (pb) que se constituyen al enfrentarse las cadenas no lo hacen al azar, sino que siempre se aparean una base púrica y una pirimídica; más específicamente: adenina con timina y citosina con guanina.
Este apareamiento específico se produce por dos motivos: por una limitación estérica (espacial) y por la composición de las bases. La limitación estérica se debe a las bases púricas son más grandes que las pirimídicas (7 y 4 Aº, respectivamente). Si las bases se enfrentasen al azar, la molécula de ADN no tendría un diámetro constante, sino zonas anchas, donde se acoplaran dos base púricas, angostas, donde se unieran dos pirimídicas, e intermedias, si el par se formara con una base de cada tipo. Al complementarse una base púrica con una pirimídica, el ancho del par de bases y de la molécula toda, es constante (par de bases=7+4=11). Esto hace que la estructura sea más estable.
Además, tanto la adenina y la timina por un lado, como la citosina y la guanina por el otro, poseen sustituyentes en las posiciones adecuadas para establecer uniones puentes de hidrógeno. Entre adenina y timina se forman dos y entre citosina y guanina, tres puentes de hidrógeno.
Esta regla de complementaridad entre las bases determina que la composición de bases de una cadena quede absolutamente supeditada a la de la otra cadena que forma la molécula; ambas cadenas son complementarias. Como ya veremos, este hecho es esencial para los mecanismos de autoduplicación y transcripción.
En lo que respecta a la secuencia de bases de una cadena, no existen restricciones: las bases pueden sucederse en cualquier orden.
 |
Las dos cadenas antiparalelas y complementarias adoptan en el espacio una estructura helicoidal dextrógira (con giro hacia la derecha), como si se enrollaran sobre un cilindro imaginario. Por eso se ha comparado a la molécula de ADN con una escalera de caracol. Las barandas estarían representadas por los ejes laterales de las cadenas, sucesiones de pentosa y fosfato unidos por los puentes fosfodiéster. Entre ambas “barandas”, los pares de bases, perpendiculares al eje, serían los escalones. Este tipo de disposición, en la cual pentosas y fosfatos se exponen en el exterior, mientras las bases se ocultan en el interior de la doble hélice, responde al comportamiento de dichas moléculas en medio acuoso. En efecto, la pentosa y el fosfato son hidrofílicos, en tanto las bases son hidrofóbicas.
El ancho total de la molécula de ADN es de 20 Aº; los pares de bases o “escalones”, se suceden a intervalos de 3,4 Aº a lo largo de la molécula. Debido al giro de la molécula, cada par de bases tiene una rotación de 36º con respecto al anterior. Esto significa que, cada 34 Aº, la molécula abarca 10 pb y completa un giro de 360º.
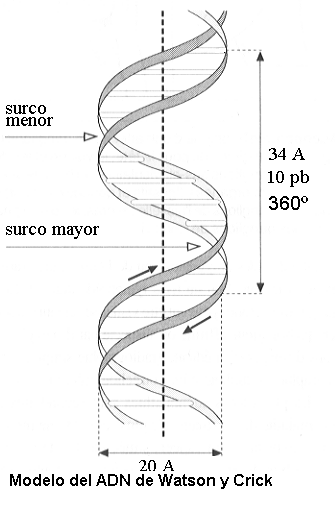 |
La descripción precedente de la estructura del ADN corresponde a un modelo presentado conjuntamente en 1953 por el biólogo James Watson y el físico Francis Crick, el cual les valió el Premio Nobel de Fisiología o Medicina, en 1962; se conoce como “el modelo de Watson y Crick”.
Watson y Crick dilucidaron la estructura de ADN partiendo de algunos datos previos, que lograron unir, literalmente, como piezas de un rompecabezas. Hasta entonces, se conocía la composición química del ADN, pero no la forma en que sus componentes se unen. Chargaff ya había señalado, unos años antes, que en el ADN de distintas procedencias se cumplían las siguientes reglas: el porcentaje de adenina coincidía con el de timina y el porcentaje de guanina coincidía con el de citosina.
Por otro lado, estudios cristalográficos realizados sobre el ADN con la técnica de difracción de rayos X, ofrecían imágenes del ADN que parecían compatibles con una estructura helicoidal. Estos datos habían sido obtenidos por Rosalind Franklin y Maurice Wilkins, quienes los pusieron en conocimiento de Watson y Crick mientras desarrollaban su investigación.
Watson y Crick construyeron modelos moleculares tridimensionales de los bloques químicos que componen el ADN. Con ellos, intentaron armar una estructura molecular tridimensional, que fuera coherente con las propiedades químicas, el comportamiento frente al agua y la geometría de dichos bloques y también con los datos previos. Así surgió el modelo del ADN. La comprensión de la estructura de esta molécula, a la cual ya se le había adjudicado el rol de material genético, fue el desencadenante del acelerado crecimiento de la Biología Molecular que se produjo desde entonces.
 |
Fórmulas de Nucleótidos y Ácidos nucleicos Regresar
Genoma y genes Regresar
El genoma de una célula es la totalidad de la información genética contenida en su secuencia completa de ADN. (Cabe aclarar que al hablar de “secuencia”, nos referimos a la secuencia de nucleótidos. Sin embargo, como éstos solamente se diferencian en la base, se utilizan las expresiones “secuencia de nucleótidos” y “secuencia de bases” en forma indistinta).
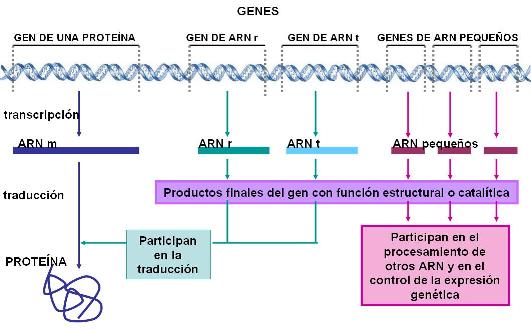
Algunas partes en la secuencia de un genoma son genes; otras partes son regiones intergénicas.
- Entre los genes se incluyen los fragmentos que contienen información para fabricar una proteína. Los genes que llevan información codificada para producir proteínas se transcriben en ARNm. Esto es, se fabrica una molécula de ARNm que copia la secuencia de bases del gen. Luego, la información que lleva codificada el ARNm es decodificada o traducida en los ribosomas. Como resultado, se fabrica una proteína según las instrucciones del gen.
- Otros genes se transcriben, pero no se traducen. Significa que un fragmento de ADN se utiliza como molde para sintetizar una molécula de ARN, pero éste no tiene información codificada para fabricar una proteína. El ARN que se obtiene en este caso es el producto final del gen y tiene otra función, que puede ser estructural, o catalítica. En alguna de estas dos categorías entran todos los demás tipos de ARN, distintos del ARNm.
Por lo tanto, un gen puede definirse como un fragmento dentro de la secuencia de ADN, que codifica una proteína o una molécula de ARN estructural o una molécula de ARN catalítico.
 |
Sin embargo, dentro de los genes hay regiones señalizadoras, que no se transcriben. Éstas actúan como signos de puntuación, marcando el principio y el final de un gen. Si bien estas zonas no son copiadas, son indispensables para el que el resto del gen se transcriba. Entonces podría ampliarse la definición de gen, incluyendo no solo a las regiones que codifican una proteína o un ARN, sino también a las secuencias requeridas para transcribirlo. El gen es una “unidad de transcripción”.
Las células regulan la expresión de sus genes; no transcriben y traducen todos sus genes al mismo tiempo. Existen en el genoma secuencias que actúan como reguladoras. Son regiones no codificantes, pues no llevan información para fabricar otras moléculas ni se transcriben, pero cumplen la función de regular el momento y la velocidad de transcripción de los genes.
Es decir que el ADN no sólo contiene la información sobre todas las proteínas y ARN, sino que también, por medio de las secuencias reguladoras, informa acerca de cuándo, en qué células y en qué cantidad se deben fabricar. Para ejercer la regulación de la transcripción, las secuencias reguladoras deben unirse con proteínas reguladoras, a las que se conoce como factores de transcripción específicos.
Las secuencias intergénicas son secuencias no codificantes. Nunca se transcriben y aunque existen diversas hipótesis sobre su significado, se desconoce su función. Se las ha llamado “ADN chatarra” o “ADN basura” o “exceso de ADN”. Muchas de ellas son elementos transponibles o móviles, secuencias que pueden cambiar de posición dentro del genoma. En el genoma humano representan alrededor del 50% de la secuencia total.
Debe quedar claro que el ADN no es otra cosa que un polímero de nucleótidos. Las regiones del ADN, codificantes, no codificantes, señalizadoras y reguladoras, no están separadas por ningún límite físico, solamente se diferencian entre sí por la secuencia de las bases que componen los nucleótidos. La información genética está codificada en la secuencia de bases.
En el año 2001 se publicó el primer esbozo completo de la secuenciación llevada a cabo por el Proyecto Genoma Humano. El genoma humano consta de un total 3,2 x 109 pb y alrededor de 30.000 genes.
Transcripción Regresar
La transcripción es el proceso en el cual la secuencia de bases de un gen se utiliza como molde para la síntesis de una molécula de ARN.
Cada gen presenta una región promotora o promotor (una secuencia de bases específica) que señaliza el sitio donde comienza el gen y el nucleótido de la secuencia donde debe iniciarse la transcripción. A su vez, otras secuencias terminadoras del gen marcan el punto final de la transcripción.
La transcripción de un gen requiere la actuación de una enzima ARN polimerasa (ARN pol). Ésta reconoce la secuencia del promotor y se une a él. En eucariotas, la unión de las ARN polimerasas al promotor está mediada por un grupo de proteínas llamadas factores basales de transcripción.
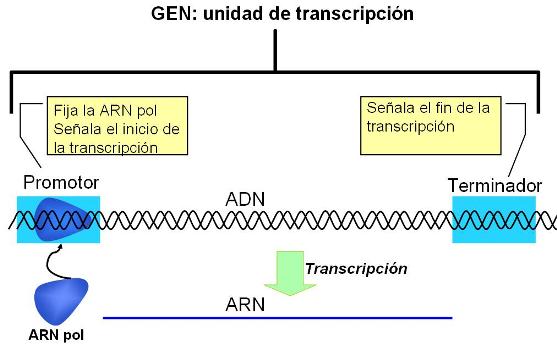 |
A partir de su unión con el promotor, la ARN polimerasa avanza sobre el gen, separando transitoriamente las dos cadenas complementarias de ADN que lo forman. Las cadenas o hebras de ADN desapareadas forman una “burbuja de transcripción”. Una de las dos cadenas, la que la ARN polimerasa recorre de 3’ a 5’, es utilizada como molde o plantilla para ser copiada. La cadena complementaria, que queda orientada de 5’ a 3’, es el antimolde de ese gen.
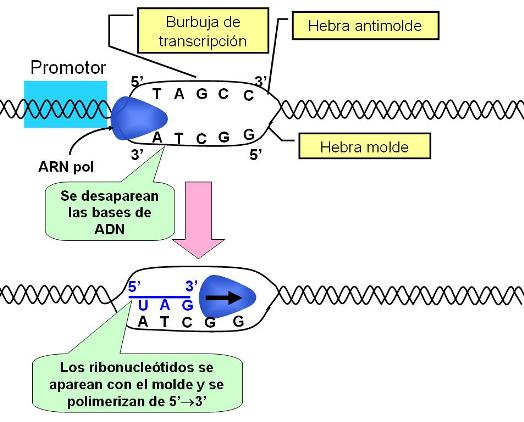 |
A medida que la ARN pol avanza, reconoce las bases que forman la cadena molde. Sobre cada nucleótido del molde la ARN pol ubica un ribonucleótido con la base complementaria. (Los ribonucleótidos se aparean con los desoxirribonucleótidos según las mismas reglas de complementaridad que rigen para el ADN, con la sola excepción de que la timina es reemplazada por uracilo.) Conforme los ribonucleótidos se aparean con el molde de ADN, la ARN pol cataliza la formación de los puentes fosfodiéster que los enlazan entre sí. De esta forma va creciendo una cadena de ARN complementaria y antiparalela al molde. El ARN crece en la dirección 5’->3’ sobre un molde “leído” en la dirección 3’->5’.
Al mismo tiempo que el transcripto de ARN crece, la burbuja de transcripción se desplaza junto con la polimerasa. La parte del molde ya transcripta vuelve a aparearse con el antimolde y el extremo 5’ del transcripto se separa del molde.
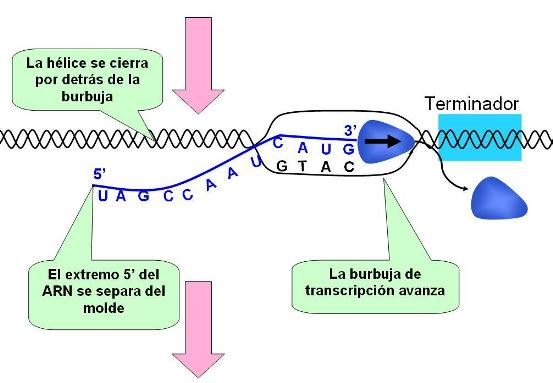 |
El proceso concluye cuando la ARN pol llega a la secuencia terminadora. Entonces el extremo 3’ del ARN sintetizado se desaparea de su molde y se libera, mientras la burbuja de transcripción se cierra. Como el ARN se sintetiza por complementaridad con respecto al molde y antiparalelo al mismo, su dirección y secuencia de bases coinciden con las del antimolde (excepto porque donde éste tiene timina, aquél tiene uracilo). Por esta razón, el antimolde es también llamado hebra positiva o codificante; el molde es la hebra negativa o no codificante.
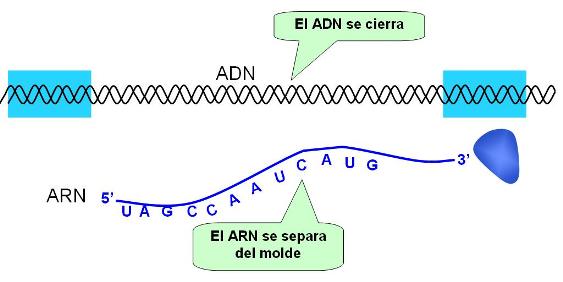 |
En los eucariotas la transcripción se lleva a cabo en el núcleo celular. Allí se localizan las ARN polimerasas, los factores basales de transcripción, los ribonucleótidos trifosfato que actúan como precursores o materia prima para fabricar el ARN, y otras enzimas que forman parte de la “maquinaria transcripcional”.
En los eucariotas existen tres clases de ARN polimerasas, con sus respectivos factores basales. Las ARN polimerasas reconocen específicamente a determinado tipo de promotores y se especializan, por lo tanto, en la transcripción de genes de diferentes ARN.
Código genético Regresar
En la secuencia de bases de los ARNm está codificada la información para la síntesis de proteínas. Un gen de una proteína X tiene una determinada secuencia de bases, ésta se transcribe al ARNm y en ella debe radicar la información acerca de cuáles aminoácidos, cuántos y en qué orden deben ser unidos para sintetizar la proteína X.
¿Cómo es que una secuencia de nucleótidos puede informar acerca de una secuencia de aminoácidos?
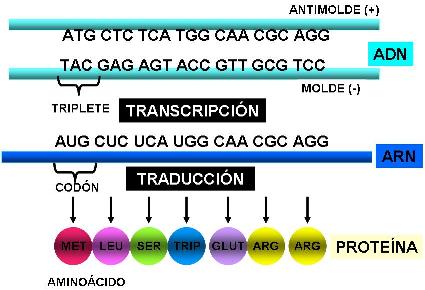
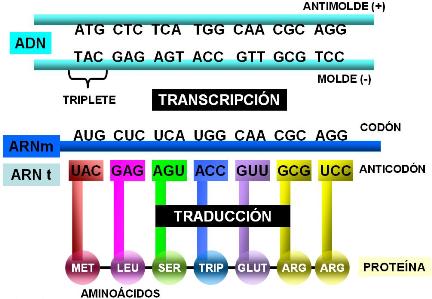
El código o idioma de los genes está constituido por tripletes de nucleótidos. Las cuatro bases (adenina, guanina, citosina y timina o uracilo) pueden formar 64 tripletes diferentes, según el orden en que se combinen. Cada triplete funciona como una palabra del código genético. Los tripletes, una vez transcriptos al ARNm, reciben el nombre de codones (codón: unidad del código).
De los 64 tripletes o codones existentes, 3 son interpretados como punto final del mensaje; son los codones stop: UGA, UAG Y UAA. Cuando el ARNm es traducido en los ribosomas, cualquiera de los codones stop indica que la proteína ya ha sido terminada.
Los 61 codones restantes codifican aminoácidos. Cada codón nombra a uno y solo un aminoácido particular. Esto evita errores en el momento de la traducción. Si un codón nombrara a dos aminoácidos diferentes, la célula no sabría cuál de ellos escoger cada vez que se encontrara ese codón. O sea que el mensaje sería ambiguo, pues daría lugar a dos interpretaciones distintas. Pero el código genético tiene, para cada codón, una interpretación única: no es ambiguo.
Ahora bien, existen 61 codones para codificar aminoácidos y los aminoácidos son solo 20. ¿Cuál es la función de los codones restantes? Los codones restantes funcionan como sinónimos. Es decir que cada aminoácido está codificado por más de un codón, con la excepción de metionina y triptófano que están nombrados por un único codón cada uno. La existencia de diferentes codones sinónimos que codifican a un mismo aminoácido es una característica del código que se conoce como degeneración o redundancia.
SEGUNDA BASE |
|
U |
C |
A |
G |
|
PRIMERA BASE |
U |
UUU fenilalanina |
UCU serina |
UAU tirosina |
UGU cisteína |
U |
TERCERA BASE |
| UUC fenilalanina |
UCC serina |
UAC tirosina |
UGC cisteína |
C |
| UUA leucina |
UCA serina |
UAA stop |
UGA stop |
A |
| UUG leucina |
UCG serina |
UAG stop |
UGG triptófano |
G |
| C |
CUU leucina |
CCU prolina |
CAU histidina |
CGU arginina |
U |
| CUC leucina |
CCC prolina |
CAC histidina |
CGC arginina |
C |
| CUA leucina |
CCA prolina |
CAA glutamina |
CGA arginina |
A |
| CUG leucina |
CCG prolina |
CAG glutamina |
CGG arginina |
G |
| A |
AUU isoleucina |
ACU treonina |
AAU asparagina |
AGU serina |
U |
| AUC isoleucina |
ACC treonina |
AAC asparagina |
AGC serina |
C |
| AUA isoleucina |
ACA treonina |
AAA lisina |
AGA arginina |
A |
| AUG metionina |
ACG treonina |
AAG lisina |
AGG arginina |
G |
| G |
GUU valina |
GCU alanina |
GAU aspartato |
GGU glicina |
U |
| GUC valina |
GCC alanina |
GAC aspartato |
GGC glicina |
C |
| GUA valina |
GCA alanina |
GAA glutamato |
GGA glicina |
A |
| GUG valina |
GCG alanina |
GAG glutamato |
GGG glicina |
G |
Durante la traducción, el mensaje que porta el ARN se lee de corrido. El primer codón AUG (codifica al aminoácido metionina) que aparece en el ARNm, es tomado como el primer codón del mensaje o codón de iniciación. De allí en más, el mensaje es traducido secuencialmente (cada tres nucleótidos: un codón), sin que existan nucleótidos sin traducir y sin alterar el orden.
 |
La característica más significativa del código genético es su universalidad. El código genético es universal: es el mismo para todos los seres vivos. Esto significa que un gen humano puede ser introducido en una bacteria y que la bacteria puede traducirlo, fabricando la misma proteína que fabricaría la célula humana.
La universalidad del código es lo que ha permitido la creación de organismos “transgénicos”. Un organismo transgénico es aquél en el cual se introducen genes provenientes de otra especie, con el fin de que fabrique determinadas proteínas. Los transgénicos se utilizan en la investigación, en la industria farmacéutica y también en la producción animal y vegetal, para dotar a plantas y animales de características que mejoran su calidad o rendimiento.
Pero más allá de las aplicaciones prácticas que ha posibilitado, la universalidad del código genético es una prueba más y muy contundente de que los seres vivos evolucionamos a partir de un único ancestro. Es muy poco probable que el mismo código hubiera surgido repetidas veces en forma independiente. La explicación de que todos compartamos un código universal no puede ser otra que la del origen común de todos los seres vivos.
ARN mensajero (ARNm) Regresar
El ARNm es la molécula que lleva la información para la síntesis de una proteína. Durante muchos años se sostuvo que el gen, el ARNm y la proteína son “colineales”, pues la secuencia de nucleótidos del mensaje se correspondería con la secuencia de aminoácidos de la proteína. Esto resultó ser una verdad a medias. En la década de 1970 se encontró que los ARN recién transcriptos de los eucariotas, también llamados pre-ARNm o transcriptos primarios, suelen ser mucho más largos mientras permanecen en el núcleo, que sus versiones maduras, ya exportadas al citoplasma. Esto se debe a que en el gen existen secuencias que se transcriben, pero que luego se eliminan del ARN. En efecto, en los genes que codifican proteínas en los eucariotas, se presentan dos tipos de secuencias que se alternan: los exones y los intrones. Tanto exones como intrones son transcriptos. Sin embargo, una vez formado el transcripto primario, los intrones son escindidos y se eliminan del ARNm, mientras que los exones se empalman unos con otros. Este proceso recibe el nombre de corte (de intrones) y empalme (de exones) o “splicing”.
En el núcleo de las células eucariotas existen partículas ribonucleoproteicas, los espliceosomas, formados por ARN pequeño nuclear (ARNsn =”small nuclear”) y proteínas, cuyas moléculas de ARN catalizan el corte de intrones y el empalme de exones.
La palabra exón significa “que se expresa”, pues los exones son las partes traducidas; los intrones son las regiones “intercaladas”. El mensaje para la síntesis de la proteína queda constituido una vez que los exones se empalman. No obstante, la secuencia de codones queda flanqueada, tanto en el extremo 5’ como en el 3’, por regiones no codificantes. Se trata de secuencias necesarias para la traducción, pero que no se traducen en aminoácidos.
Además del corte y empalme, los pre-ARNm eucariotas sufren otras dos modificaciones antes de convertirse en ARNm maduros aptos para ser traducidos. La primera de ellas es el “capping”. Consiste en el agregado de un “capuchón” que no es otra cosa que un nucleótido modificado, en el extremo 5’ del transcripto. El capuchón o “cap” se añade al transcripto cuando éste consta de unos pocos nucleótidos de longitud, mientras aún se está transcribiendo. La cap protege al ARN de enzimas que pueden degradarlo y es necesario para el splicing y la posterior traducción.
Por último, en el extremo 3’ del transcripto, se realiza la poliadenilación. Es el agregado de una serie de nucleótidos de adenina que forman la cola poli-A. Aunque existen excepciones, en general la cola poli-A parece ser indispensable para que el ARNm pueda salir por los poros de la envoltura nuclear.
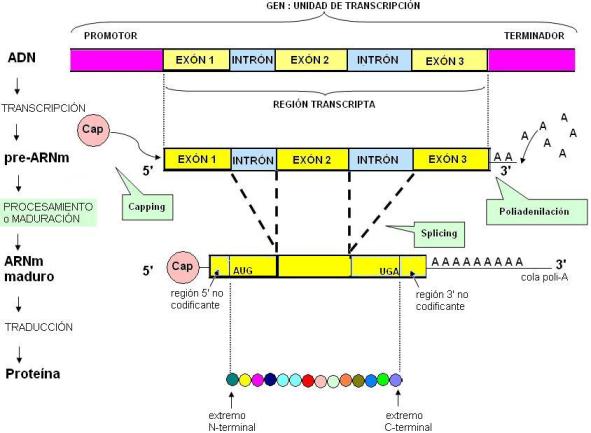 |
De todo lo dicho se desprende que el gen es realidad más largo que el transcripto, y éste más largo que el ARNm. Además, en el ARNm maduro aparecen nucleótidos que no son copiados del gen (la cap y la cola poli-A) y regiones que no serán traducidas.
Los ARNm, una vez maduros o procesados, se dirigen al citosol, donde son traducidos (aunque descubrimientos recientes sugieren que también podría haber traducción en el núcleo). Después de la traducción, en general son rápidamente degradados. Esta rápida degradación es una forma de regular la cantidad de proteína sintetizada.
ARN de transferencia (ARNt) Regresar
Los ARNt son moléculas que se encargan de transportar los aminoácidos, es decir, la materia prima para la síntesis de una proteína, hasta el ribosoma.
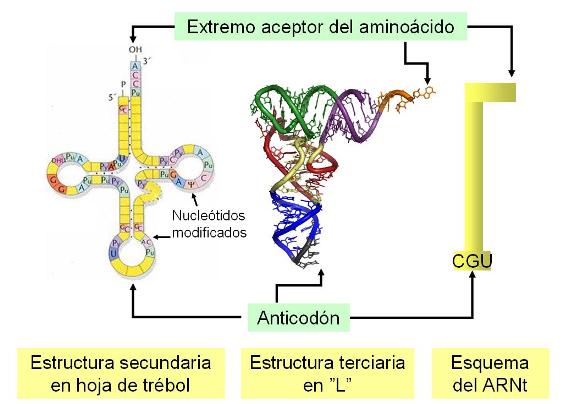
Cada ARNt es una molécula lineal de unos 75 - 85 nucleótidos de longitud, que se pliega adoptando una estructura secundaria en forma de hoja de trébol. Un nuevo plegamiento, sobre la estructura secundaria, da lugar a una estructura terciaria en forma de “L”.
En los ARNt ya plegados se distinguen dos extremos: el extremo aceptor del aminoácido y el anticodón. El extremo aceptor se une a un aminoácido determinado. El anticodón consiste en una secuencia de tres bases de la cadena del ARNt, la que se unirá a un codón que tenga una secuencia complementaria.
 |
Los ARNt son específicos para una clase de aminoácido; por otro lado, su anticodón reconoce sólo al codón que lo complementa. Como resultado, a cada codón le corresponde un aminoácido específico. Puede decirse que los ARNt son los verdaderos “traductores”, ya que al mismo tiempo son capaces de leer el idioma de los nucleótidos en el ARNm y de reconocer a los aminoácidos, haciendo de enlace entre ambos lenguajes.
 |
La unión de cada ARNt con el aminoácido “correcto” depende de la actividad de unas enzimas llamadas aminoacil-ARNt sintetasas. Cada una de ellas cataliza la formación del complejo entre el aminoácido y el ARNt específico (complejo aminoacil-ARNt). Las aminoacil-ARNt sintetasas reconocen algunas bases que diferencian a un ARNt de otro. La reacción en la cual se produce la unión se denomina activación y consume energía. La energía queda parcialmente retenida en el enlace de alta energía que se forma entre el ARNt y el aminoácido y será utilizada para la formación del enlace peptídico durante la traducción. La activación tiene lugar en el citosol, antes de la traducción propiamente dicha.
Dado que todas las proteínas se construyen a partir de los mismos veinte aminoácidos, cada ARNt puede utilizarse para traducir cualquier mensaje. Así es que, a diferencia de lo que ocurre con los ARNm, los ARNt son moléculas más estables, que tienen una vida media larga y se reutilizan una y otra vez.
ARNr y ribosomas Regresar
El ARN ribosomal es el ARN más abundante en las células. En eucariotas existen cuatro tipos de ARN ribosomales (ARNr). De ellos, tres se transcriben en el nucléolo como un solo ARN precursor, que luego es escindido. Al cortarse el transcripto precursor, se obtienen los tres ARNr, de menor tamaño. El cuarto ARNr se transcribe fuera del nucléolo.
Los distintos tipos de ARN ribosomal se asocian con proteínas para formar las subunidades ribosomales. La construcción de las subunidades también se produce dentro del nucléolo.
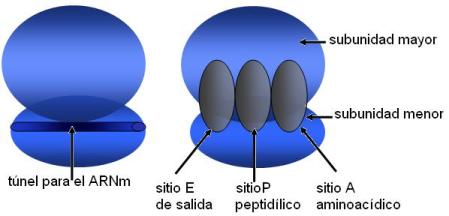
La función de los ribosomas es la síntesis de proteínas. Los ribosomas se ensamblan en el citoplasma, cuando inician la traducción de un ARNm. En un ribosoma funcional quedan delimitados dos canales: uno en la subunidad menor, donde se ubica el ARNm, y otro en la subunidad mayor, por donde emerge la proteína en síntesis.
Internamente, la estructura del ribosoma posee tres cavidades, denominadas sitios A, P y E, que abarcan tanto la subunidad menor como la mayor.
El sitio A o aminoacídico aloja a los ARNt, que ingresan al ribosoma cargados con sus respectivos aminoácidos para ser incorporados a la proteína.
El sitio P o peptidílico es el sitio donde crece la cadena peptídica que se está sintetizando.
En el sitio E (por “exit”) se ubican los ARNt prontos a abandonar el ribosoma, después de que el aminoácido que portaban se haya unido a la proteína en fabricación.
 |
Traducción Regresar
La traducción se inicia al unirse el ARNm con el ARNt iniciador, que transporta metionina (met), y las subunidades ribosomales. El ribosoma se ensambla en el extremo 5’ del ARNm y se ubica de modo tal que el primer codón AUG desde el extremo 5’, codón de iniciación, quede situado en el sitio P. Sobre el codón de iniciación se coloca el ARNt iniciador cargado con metionina.
| Etapa de iniciación |
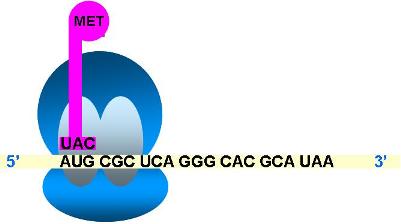 |
Se asocian:
- subunidad menor
- ARNt iniciador
- ARNm
- subunidad mayor
|
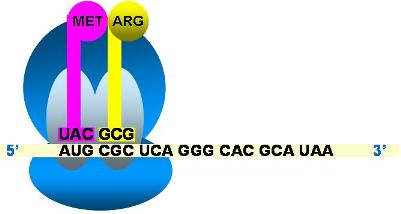
Una vez completada esta primera etapa, llamada etapa de iniciación, comienza la elongación de la cadena peptídica. Entonces un segundo ARNt, cuyo anticodón complementa al codón adyacente a AUG, en sentido 3’, ingresa al sitio A, cargando el segundo aminoácido. A continuación, el primer aminoácido se separa de su ARNt y forma enlace peptídico con el segundo aminoácido, aún unido a su propio ARNt. Ahora, el segundo ARNt, ubicado en el sitio A, lleva un dipéptido. El ribosoma se desplaza sobre el ARNm en sentido 5’->3’ (translocación). El ARNt con el péptido pasa del sitio A al sitio P, dejando vacante al primero. Al mismo tiempo, el ARNt iniciador ingresa al sitio E antes de salir del ribosoma.
| Etapa de elongación |
 |
-
Ingresa al sitio A el ARNt complementario del segundo codón.
-
Los sitios P y A están ocupados por sendos ARNt con sus respectivos aminoácidos.´
-
La peptidil transferasa cataliza la formación del enlace peptídico entre los dos aminoácidos.
|
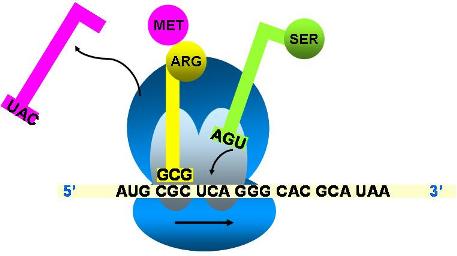 |
- Se transfiere el primer aminoácido al segundo y se libera el primer ARNt
- El ribosoma se transloca, dejando vacante el sitio A.
- Un nuevo ARNt cargado con su aminoácido ingresa al sitio A.
- El péptido permanece unido al ARNt ubicado en el sitio P.
- El péptido será transferido al aminoácido en el sitio A.
|
Durante la elongación se repite este ciclo de eventos, y a medida que el ribosoma se transloca sobre el ARNm hacia el extremo 3’, la cadena peptídica va creciendo en el sitio P y empezará a asomar por el túnel de la subunidad mayor. El primer aminoácido ingresado a la proteína lleva el extremo N-terminal; por lo tanto, éste será el primero en aparecer. El último portará el extremo C-terminal.
| Terminación de la traducción |
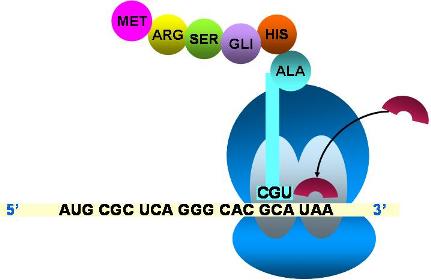 |
- El codón stop queda ubicado en el sitio A.
- Un factor de liberación se une al codón stop .
|
El fin de la traducción se produce cuando un codón stop o de paro llega al sitio A. Los codones de terminación no tienen ARNt complementarios; a ellos se unen proteínas denominadas factores de liberación. El factor de liberación provoca la separación de todos los elementos que participaron en la traducción: se separan las subunidades ribosomales del ARNm y el último ARNt rompe su unión con la proteína sintetizada.
| Terminación de la traducción |
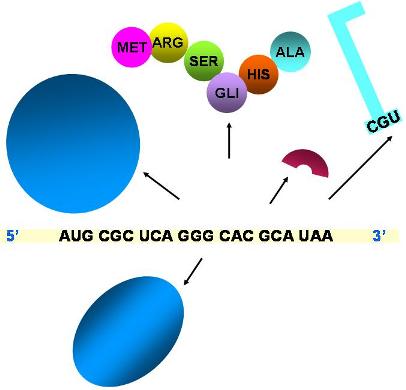 |
- Todos los elementos que participan en la traducción se separan.
- Se obtiene la proteína.
|
Es importante destacar que el ARN ribosomal cumple un papel catalítico durante la traducción. En efecto, la formación del enlace peptídico se debe a la actividad del ARNr de la subunidad mayor, que se comporta como enzima “peptidil-transferasa”. Las proteínas del ribosoma, en cambio, parecen tener un papel estructural.
Además de los factores de liberación, que actúan en la etapa de finalización, para la síntesis proteica se requiere la presencia de otras proteínas que participan en las etapas precedentes: los factores de iniciación y de elongación.
Por último, se debe recalcar que la síntesis proteica es un proceso que consume energía; fabricar proteínas es, para la célula, un proceso energéticamente más costoso que la síntesis de cualquier otro tipo de molécula.
Regulación genética: transcriptoma y proteoma Regresar
Todas las células somáticas o corporales de un mismo organismo contienen la misma información genética. Sin embargo, una neurona se distingue perfectamente de una célula muscular o de un glóbulo blanco. Estas diferencias morfológicas y funcionales se deben a que en los distintos tipos celulares y aun en la misma célula en distintas etapas de su vida, cambia la forma en que se expresa el genoma.
Las células poseen mecanismos para regular la expresión del genoma en cada uno de los pasos involucrados. Así, controlan:
- qué genes se transcriben en cada momento,
- a qué velocidad se transcriben
- qué transcriptos son procesados para que puedan llegar a traducirse,
- de qué forma son procesados dichos transcriptos,
- cuáles de los ARNm atraviesan los poros nucleares para llegar al citosol,
- qué vida media tendrá cada ARNm ya en el citoplasma,
- cuál ARNm será traducido, y a qué ritmo,
- cuál será la vida media de la proteína obtenida, y
- en algunos casos, si la proteína estará activa o no.
De todos los pasos de control mencionados, posiblemente el de mayor importancia sea el que se ejerce a nivel de la transcripción.
En síntesis: las diferencias entre células de un mismo organismo no se deben a diferencias en el genoma, sino que deben atribuirse a diferencias en su transcriptoma y su proteoma. El transcriptoma es el conjunto de todos los ARN que posee una célula en un momento dado. El proteoma es el conjunto de las proteínas presentes en determinado momento de la vida celular.
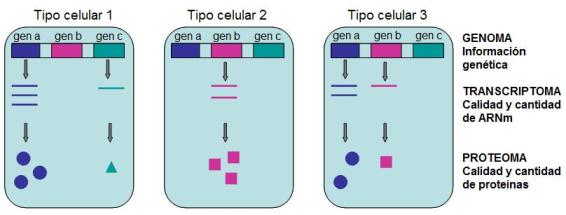 |
Las diferencias entre los distintos tipos celulares de un organismo son debidas fundamentalmente a una transcripción diferencial de sus genes, es decir a la presencia de un transcriptoma y, en consecuencia, de un proteoma diferentes. |
ADN y Ciclo celular Regresar
Ciclo celular Regresar
Las células surgen por división celular a partir de otra célula preexistente. La célula madre duplica su material genético y luego lo distribuye, de manera que cada futura célula hija reciba una copia completa del mismo. Finalmente se divide el citoplasma de la célula madre, con sus correspondientes organoides, dando lugar a la formación de dos células hijas.
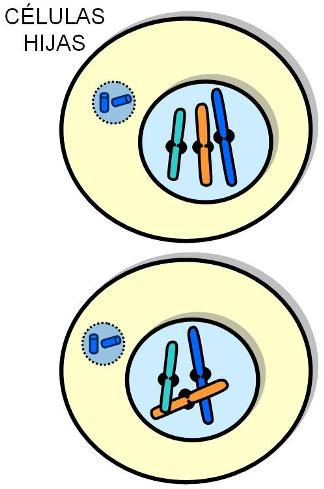
Las células hijas heredan una copia íntegra de la información genética presente en la célula original. Esto las hace potencialmente idénticas; de hecho lo son desde el punto de vista genético. Sin embargo, cada célula hija recibe aproximadamente la mitad de la masa citoplasmática original; deberá pasar entonces por un período de crecimiento hasta estar en condiciones de entrar, a su vez, en la etapa de división.
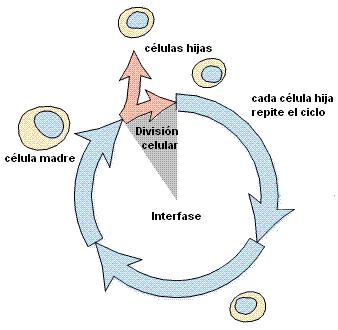 |
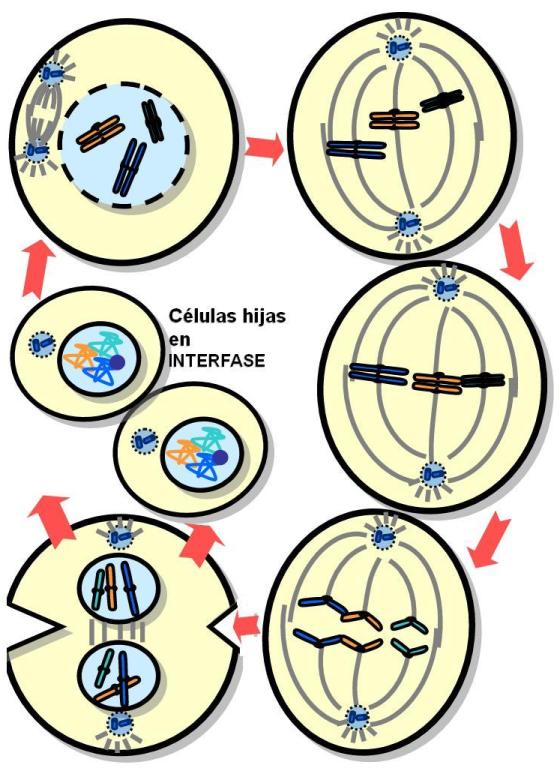
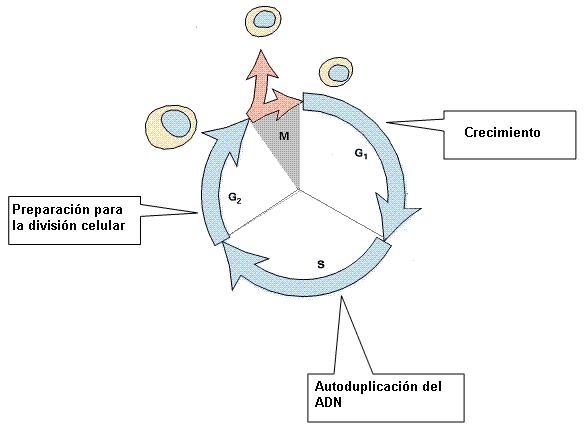
El ciclo de vida de una célula eucariota, o ciclo celular, comprende una etapa de interfase y una etapa de división celular o etapa “M”.
La interfase se subdivide en tres etapas:
-G1 (G proviene de “gap”: intervalo): ésta es la etapa de crecimiento celular. Las células sintetizan nuevos componentes, hasta alcanzar el tamaño de la célula madre. Además del crecimiento y las funciones inherentes a su automantenimiento, en los organismos pluricelulares cada célula lleva a cabo una función específica. La etapa G1 de la interfase es de una intensa actividad celular.
-S (de “síntesis”): durante esta fase la célula duplica el material genético. Cada molécula de ADN del núcleo celular se utiliza como molde para generar dos moléculas de ADN idénticas. Este proceso se denomina replicación o autoduplicación del ADN. Las dos copias idénticas permanecen unidas hasta la división celular.
-G2: es una etapa de preparación para la división celular inminente.
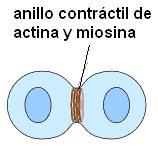
La fase M o de división celular comprende la mitosis, o división del material genético y la citocinesis o división del citoplasma.
 |
| Interfase |
G1 |
| S |
| G2 |
| Fase M |
Mitosis |
Profase |
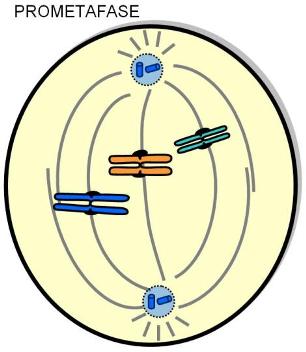
| Prometafase |
| Metafase |
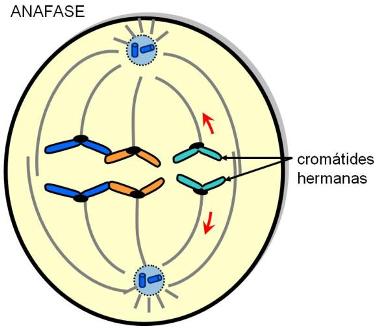
| Anafase |
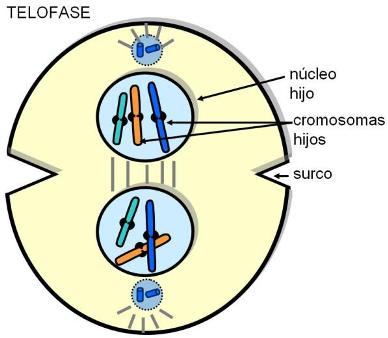
| Telofase |
| Citocinesis |
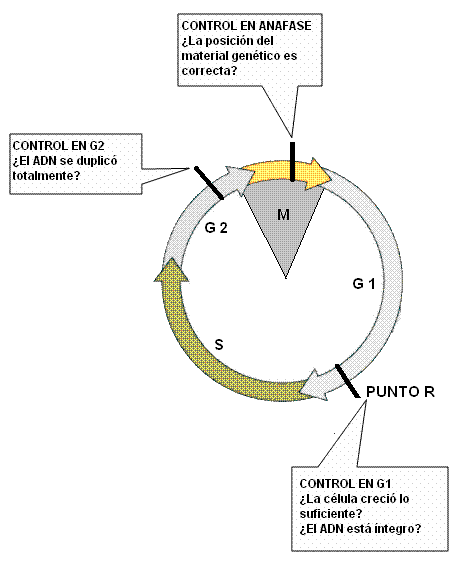
Control del ciclo celular Regresar
El ciclo celular se desarrolla según un programa que sólo permite el avance hacia otra etapa cuando la etapa en curso se ha cumplido adecuadamente. Este programa consta de algunos puntos de control cruciales. Uno de ellos es el punto R o punto de restricción, al final de la etapa G1. En este punto se chequean fundamentalmente dos variables: que el material genético no haya sufrido daño y que las condiciones del medio circundante favorezcan la división celular. Solamente si ambas variables lo permiten, entonces se produce el avance hacia la fase S.
El segundo punto de control está situado después de la fase S. En este momento, es de vital importancia chequear si la replicación del ADN se realizó en forma completa. En ese caso, la célula atraviesa el período G2 y avanza hacia la fase M.
Al promediar la fase M (en anafase) existe otro punto de control, cuya finalidad es controlar si las moléculas de ADN están ubicadas en la posición correcta, para proceder a su distribución equitativa entre las células hijas. Así se garantiza que las células hijas no reciban información de más o de menos.
 |
El control del ciclo celular está supeditado a la influencia de diferentes tipos de señales. En un organismo unicelular, como una ameba, la división celular es el mecanismo para perpetuar la especie. Entonces, el control estará influido fundamentalmente por señales que provienen del ambiente en el cual se desarrolla. Si el ambiente provee los recursos necesarios, el programa seguirá su curso: la ameba crecerá adecuadamente y eventualmente se dividirá, dando origen a dos amebas hijas. Si en cambio, el ambiente es adverso, el avance del ciclo celular se detendrá en respuesta a las señales externas.
En los organismos multicelulares la situación es otra. Cada célula debe servir a las necesidades del organismo total. Su ciclo, entonces, estará controlado por señales que provienen de otras células. Dichas señales permitirán que el programa del ciclo avance, o bien provocarán una interrupción del mismo, de acuerdo a las circunstancias. De esa forma, el ciclo celular de cada célula individual se ajusta a los requerimientos del organismo como un todo.
Ya en el medio intracelular, el avance del ciclo está mediado por complejos formados por dos tipos de proteínas: las ciclinas y las quinasas dependientes de ciclinas (cdk). Las ciclinas presentan concentraciones variables a lo largo del ciclo. Las quinasas, activadas por la presencia de las ciclinas, catalizan la fosforilación de diversas proteínas “blanco”. La fosforilación es un mecanismo que permite “encender” o “apagar” a las proteínas. Las proteínas blanco fosforiladas modifican su actividad, provocando los cambios celulares cíclicos.
El cáncer en sus múltiples formas se origina siempre a partir de una célula que falla en el control de su ciclo celular. La falla obedece a alteraciones o mutaciones en las secuencias de los genes que codifican las proteínas involucradas en los mecanismos de control. Las proteínas defectuosas, producto de los genes mutados, o bien no responden a las señales exteriores, o bien producen una respuesta exacerbada a las mismas. En algunos casos, dichas proteínas se activan aun en ausencia de las señales pertinentes. El resultado es que la célula cancerosa o maligna se divide en forma indiscriminada, originando una masa anormal de células, o tumor. Este comportamiento anárquico de las células cancerosas es la causa del conjunto de enfermedades que se agrupan bajo el nombre de cáncer.
Duración del ciclo celular Regresar
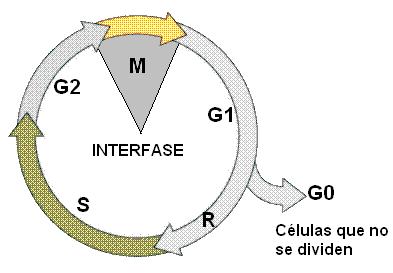
El ciclo celular tiene una duración muy variable, cuando se comparan diferentes tipos celulares. Sobre la base de este criterio, pueden reconocerse tres grandes categorías de células:
1. Células con una gran especialización estructural. Por ejemplo: neuronas o células musculares. Estos tipos celulares no se dividen; permanecen en una fase G1 que dura hasta su muerte. Como la célula no avanza hacia S, a este tipo de G1 también se la denomina G0.
2. Células que normalmente no se dividen, pero pueden hacerlo ante un estímulo apropiado. Es el caso de los hepatocitos (células hepáticas), que pasan el punto R cuando se extirpa una parte del órgano.
3. Células con gran actividad mitótica. Sus ciclos celulares se suceden con mucha celeridad. Por ejemplo, las células embrionarias o células que pertenecen a tejidos de renovación continua, como la epidermis.
La duración del ciclo celular depende fundamentalmente de la duración de la fase G1, ya que una célula no pasa el punto R si no recibe estímulos para dividirse. El resto de las fases del ciclo celular, S, G2 y fase M, duran unas pocas horas, y su duración es similar en todas las células de una especie.
 |
Estructura de la cromatina en las distintas etapas del ciclo celular Regresar
El núcleo contiene los cromosomas de la célula. Cada cromosoma consiste en una molécula única de ADN con una cantidad equivalente de proteínas. Colectivamente, el ADN con sus proteínas asociadas se denomina cromatina. La mayor parte de las proteínas de la cromatina consisten en copias múltiples de cinco clases de histonas.
Estas proteínas básicas son ricas en residuos de arginina y lisina cargados positivamente. Por esta razón se unen estrechamente con los grupos fosfatos (cargados negativamente) del ADN.
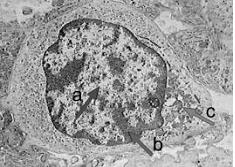
La observación de un núcleo interfásico a través del microscopio óptico nos permite distinguir dos tipos de cromatina. La eucromatina o cromatina laxa, de localización central, y la heterocromatina o cromatina densa, en la periferia del núcleo. La heterocromatina representa aproximadamente el 10% del total de cromatina y es considerada transcripcionalmente inactiva.
|
Micrografía electrónica del núcleo de un linfocito. a) eucromatina, b) heterocromatina, c) mitocondria |
Cuando el cromosoma en interfase se esparce artificialmente sobre agua, tiene la apariencia de un collar de perlas. Las perlas son los nucleosomas, las unidades de enrollamiento de la cromatina.
Los nucleosomas poseen un centro o "core" de histonas. Dicho centro posee dos copias de cada una de las siguientes histonas: H2A; H2B; H3 y H4.
Alrededor del centro de histonas, 146 pares de bases del ADN se enrollan en dos vueltas. La unión de las histonas al ADN no depende de una secuencia particular de nucleótidos, sino de la secuencia de aminoácidos de la histona. Las histonas son unas de las moléculas más conservadas durante el transcurso de la evolución. Alrededor de 60 pares de bases de ADN unen un nucleosoma con el próximo. Cada región de unión es el ADN espaciador. La quinta histona, la H1, se ubica por fuera del nucleosoma. Esta estructura se conoce como fibra de 10 nanómetros, siendo el primer grado del empaquetamiento de la cromatina.
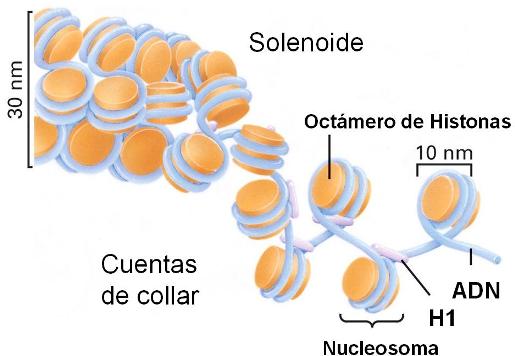 |
Los nucleosomas se organizan, a su vez, en fibras de 30 nm (solenoide), girando a manera de resorte alrededor de un eje virtual. Esta estructura es mantenida por la interacción de las H1 de nucleosomas cercanos.
En el siguiente nivel de empaquetamiento, las fibras de 30 nm se organizan en una serie de bucles o asas superenrolladas. Estos bucles se estabilizan gracias a la interacción con las proteínas de la matriz nuclear o andamiaje nuclear (“scaffold”).
Cada bucle de cromatina representa un dominio funcional o unidad de replicación. Estos dominios tienen una extensión de ADN suficiente para acomodar varios genes de tamaño promedio. Algunos genes, sin embargo, pueden abarcar varios dominios adyacentes de un cromosoma. Cada cromosoma puede tener cien o más dominios.
El grado de condensación de los dominios de cromatina se mantiene principalmente debido a la asociación con la matriz nuclear y a proteínas asociadas. La unión entre la cromatina y la matriz se da a nivel de zonas altamente conservadas, denominadas secuencias SAR o MAR (scaffold associated regions/ matrix attachment regions). Las SAR son abundantes en la heterocromatina.
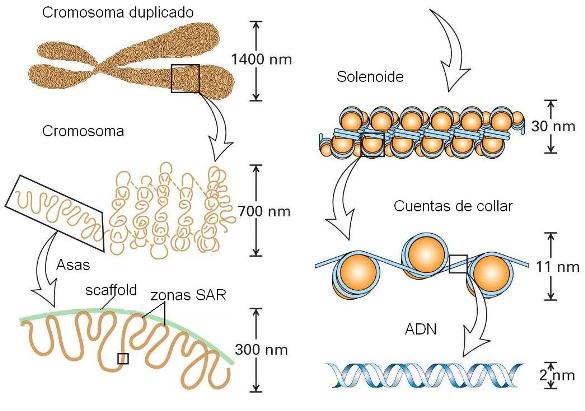 |
Al comienzo de la división celular, en la profase, los cromosomas aparecen en forma más condensada, alcanzando la cromatina su mayor nivel de condensación en metafase. El "scafffold" o matriz nuclear se convierte en el centro de la estructura del cromosoma, y como la compactación continúa, éste se pliega modo de acordeón. El empaquetamiento de la cromatina permite confinar al ADN dentro del núcleo. Midiendo extremo con extremo el total de cromosomas de una célula humana, el ADN se extiende más de 2 metros. El empaquetamiento del ADN en forma de cromatina, no solamente le permite a éste entrar dentro de los límites del núcleo, sino también lo protege del ataque de las nucleasas (enzimas que degradan ácidos nucleicos).
Estructura del cromosoma Regresar
Cada cromosoma eucariota consiste en una molécula de ADN.
La molécula de ADN en el cromosoma eucariota es lineal, por lo tanto posee dos extremos (en contraste con el cromosoma bacteriano que es circular).
La molécula de ADN de un cromosoma típico eucariota contiene un conjunto de genes y muchas secuencias de ADN no codificante.
El ADN no codificante incluye:
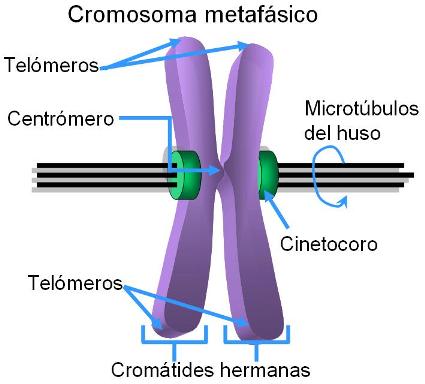
- El centrómero: está formado por secuencias de aproximadamente 170 nucleótidos, repetidas miles de veces.
- Los telómeros: secuencias repetitivas en los extremos del cromosoma.
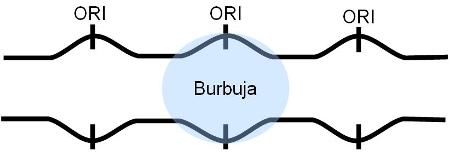
--Los orígenes de replicación (ORI): son múltiples secuencias señalizadoras altamente conservadas, necesarias para que se realice la duplicación del ADN en un tiempo breve.
El centrómero es una constricción primaria localizada centralmente o hacia los extremos de cada cromosoma.
El ADN centromérico, como ya mencionamos, es altamente repetitivo y se encuentra siempre condensado siendo parte de la heterocromatina.
Los telómeros son cruciales en la vida de la célula. Ellos son necesarios para la duplicación completa del cromosoma, los protegen de las nucleasas, evitan que los extremos del cromosoma se fusionen entre sí y facilitan la interacción del cromosoma con la envoltura nuclear.
Antes de que una célula se divida, cada cromosoma se duplica (durante la fase S del ciclo celular).
Durante la mayor parte de la vida de la célula, los cromosomas son demasiado largos y tenues para ser vistos bajo un microscopio. Al inicio de la división celular, los cromosomas duplicados se condensan en estructuras que pueden teñirse con facilidad (por ello denominadas cromosomas: cuerpos coloreados), pudiéndose observar bajo el MO.
 |
Micrografía electrónica de un cromosoma metafásico
|
La condensación es tal que el cromosoma es aproximadamente 10.000 veces más corto que la molécula de ADN que contiene. A primera vista, los cromosomas duplicados se mantienen juntos por el centrómero. Mientras están juntos, es común llamar cromátida hermana a cada parte del cromosoma duplicado.
Esto no debe confundirnos, cada una de las "cromátidas hermanas" es un cromosoma completo. El cinetocoro es una estructura proteica discoidal que forma parte del centrómero y ayuda a separar las cromátidas hermanas. Es el sitio de unión con los microtúbulos del huso, que contienen los motores de dineína que tiran a los cromosomas en la anafase. Además proveen una plataforma para ensamblar y movilizar las proteínas que construyen el huso.
Clasificación morfológica de los cromosomas Regresar
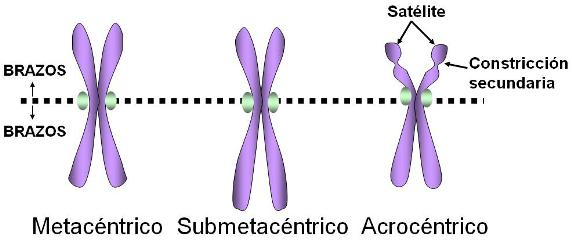
La posición del centrómero determina el largo de los brazos del cromosoma; en base a esto se puede clasificar a los cromosomas en:
1. Metacéntricos: el centrómero en posición central determina brazos de igual longitud.
2. Submetacéntricos: un par de brazos es más corto que el otro, pues el centrómero se encuentra alejado del centro.
3. Acrocéntricos: el centrómero se halla próximo a uno de los extremos, por lo tanto uno de los brazos es casi inexistente.
En células humanas, los cromosomas acrocéntricos poseen una masa de cromatina llamada satélite, en el extremo del brazo corto. El satélite se halla aislado del resto del cromosoma por la constricción secundaria. La zona aledaña al satélite de los cromosomas acrocéntricos contribuye a formar el nucléolo.
Autoduplicación del ADN Regresar
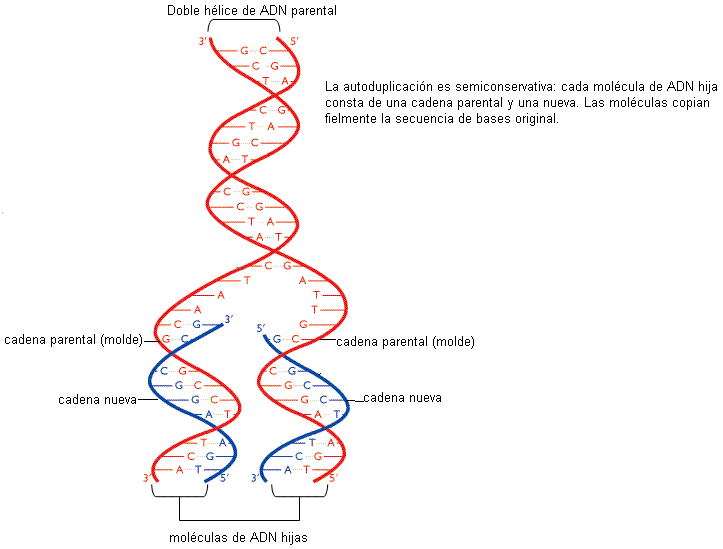
La autoduplicación o replicación del ADN es el proceso por el cual, partiendo de una molécula de ADN que se utiliza como molde, se obtienen dos moléculas de ADN idénticas.
El nombre de autoduplicación no hace referencia a que el ADN se baste a sí mismo para duplicarse; por el contrario, este proceso requiere una importante batería de enzimas y otras proteínas colaboradoras. Autoduplicación se refiere a que, debido a la estructura de la molécula, cada una de las dos cadenas puede ser usada como plantilla para la síntesis de una cadena complementaria, es decir que lleva en sí misma la información para sintetizar la copia.
Durante la autoduplicación, las dos cadenas que conforman la molécula de ADN de cada cromosoma se desaparean y sobre las bases expuestas se construye la cadena nueva. Así resultan dos moléculas de ADN. Ambas conservan una sola cadena de la molécula original o parental, mientras que la complementaria es la cadena sintetizada de novo. Se dice por esta característica que la autoduplicación es semiconservativa. Sin embargo, la secuencia de bases, donde radica la información, es copiada fielmente.
En las células eucariotas, la replicación del ADN de los cromosomas se lleva a cabo totalmente durante el período S de la interfase, en el cual el grado de plegamiento de la cromatina es menor que el observado en el período de división celular. La duplicación completa del ADN es un requisito previo para la división. La presencia de múltiples orígenes de replicación (ORI) en los cromosomas eucariotas facilita el progreso rápido del proceso, pues éste se inicia simultáneamente en todos los orígenes. Todo el ADN generado a partir de un ORI se denomina replicón. El replicón es la unidad de replicación.
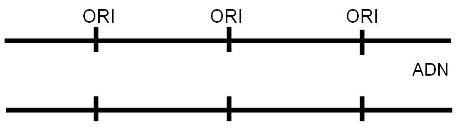 |
Los cromosomas eucariotas tienen múltiples puntos de origen de replicación (ORI). |
A la altura de los orígenes de replicación se abre la doble hélice, generándose una burbuja de replicación, que avanza progresivamente hacia ambos lados del origen, hasta encontrarse con las burbujas correspondientes a los ORI adyacentes, en los llamados puntos T o de terminación. A medida que la burbuja avanza, se sintetizan las cadenas de ADN nuevas, también en ambas direcciones. La autoduplicación es entonces bidireccional.
|
A partir de cada ORI, la doble hélice se abre progresivamente en ambas direcciones, formando una burbuja de replicación.
|
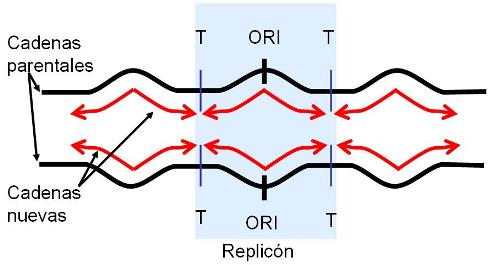 |
Las cadenas nuevas crecen bidireccionalmente a partir de cada ORI, sobre los moldes proporcionados por las cadenas parentales. |
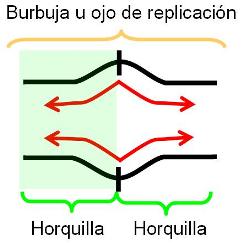 |
Las dos mitades de una burbuja se llaman horquillas. Una enzima helicasa es la encargada de abrir la doble hélice en cada horquilla. Las proteínas de unión a hebra simple (SSBP: single qtrand binding proteins) se asocian a las cadenas de ADN evitando que éstas vuelvan a aparearse. Otras enzimas, las topoisomerasas, corrigen tensiones generadas en el ADN cuando se abren los ORI. |
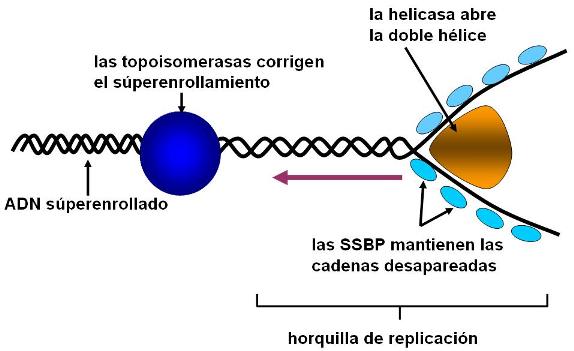 |
Las responsables de la síntesis de ADN son las enzimas de la familia de las ADN polimerasas (ADNpol). Las ADN pol tienen una serie de características que determinan, en gran medida, la forma en que se lleva adelante el proceso de replicación:
1) Las ADN pol son polimerasas 5’->3: solo sintetizan cadenas de ADN en sentido 5’->3’, recorriendo el molde en dirección 3’->5’.
2) Las ADN pol no tienen la capacidad para iniciar la síntesis de una cadena polinucleotídica; solo pueden agregar nucleótidos al extremo 3’ de una cadena ya iniciada.
3) Las ADN pol tienen actividad exonucleasa 3’->5’: significa que pueden eliminar al nucleótido que se encuentra en el extremo 3’ de la cadena. (Aunque algunas poseen además la actividad exonucleasa 5’->3, la que les permite eliminar el nucleótido del extremo 5’ de la cadena.) |
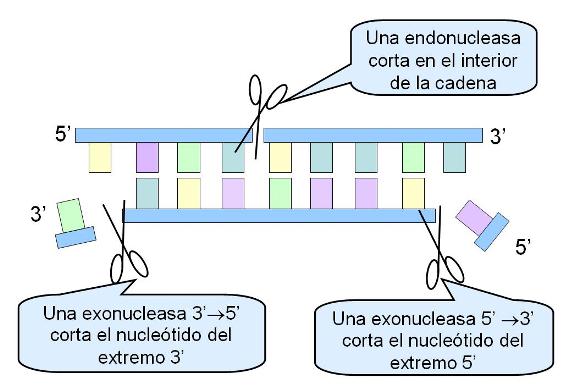 |
Consideremos el proceso de replicación en una horquilla:
Las cadenas que forman una molécula de ADN son antiparalelas. Si las dos cadenas de una horquilla han de ser copiadas a partir del origen hacia el punto de terminación, una de las cadenas nuevas tendrá que crecer en dirección 5’->3 y la otra, en la dirección opuesta, 3’->5’. Teniendo en cuenta que la actividad polimerasa de la ADN pol se cumple sólo de 5’->3’, una de las cadenas de la horquilla está bien orientada para la copia, mientras que la otra está mal orientada.
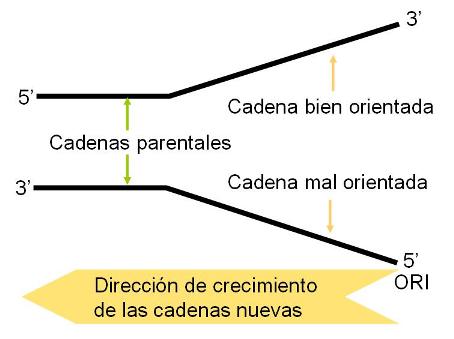 |
Sobre la cadena bien orientada, actúa en primer lugar una enzima primasa. Ésta sintetiza una corta cadena de ARN, el “primer” o cebador, que servirá para proporcionar un extremo 3’ libre a la ADN pol. La ADN pol continuará el trabajo de la primasa, agregando desoxirribonuclótidos al extremo 3’ del primer. Así fabricará una cadena de ADN, en dirección 5’->3’ y en forma continua, desde el origen a la terminación. Esta cadena crece rápidamente, por lo cual se la llama hebra líder o adelantada.
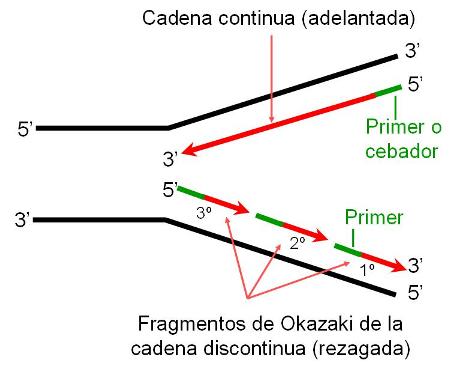 |
Sobre la cadena mal orientada, la ADN pol no empieza su trabajo exactamente en el ORI, sino que se sitúa un poco más allá del mismo, hacia el punto de terminación. Entonces, una vez que la primasa le proporciona el primer, la ADN pol se mueve sobre el molde hasta el ORI, alargando la cadena de 5’ a 3’. Cuando llega al ORI, la ADN pol se ve obligada a retroceder en dirección al punto T, a fin de sintetizar otro fragmento en dirección 5’->3’, a partir de un nuevo cebador. De esta forma se van agregando fragmentos, de manera tal que la cadena nueva parece estar creciendo desde 3’ a 5’, aunque en realidad cada fragmento fue sintetizado de 5’ a 3’, única dirección posible para la ADN pol. Los fragmentos se conocen como fragmentos de Okazaki (en el esquema se señalan como 1º, 2º, etc., por orden de síntesis).
La síntesis de ADN es discontinua ya que la cadena hija sobre la hebra mal orientada crece fragmentariamente. La cadena discontinua también se llama rezagada, pues su crecimiento es más lento que el de la cadena continua.
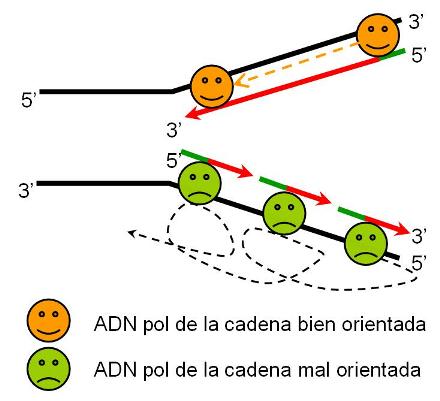 |
Cuando se amplía la mirada desde una horquilla hacia la burbuja completa, se puede observar que la cadena que resulta bien orientada en una horquilla es la que está mal orientada en la otra mitad del replicón y viceversa, ya que ambas horquillas se copian en direcciones opuestas. Es decir: no existen una cadena hija totalmente continua y otra totalmente discontinua, sino que ambas tienen sectores continuos y sectores discontinuos. A esta característica se la llama semidiscontinuidad.
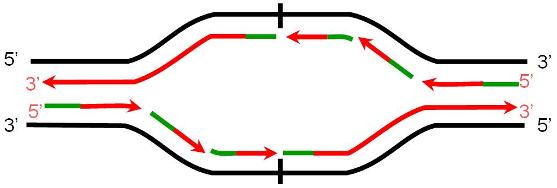 |
Semidiscontinuidad: en el replicón se observa que ambas cadena nuevas tienen partes de crecimiento continuo y partes de crecimiento discontinuo. |
Una vez completa la síntesis de las cadenas hijas, aún faltan dos trabajos por realizar. El primero de ellos es remover los primers y reemplazarlos por ADN. La remoción de los primers queda a cargo de enzimas nucleasas. El ADN que debe ocupar su lugar es sintetizado por otra enzima ADN pol. Ésta ya cuenta con extremos 3’ en los fragmentos de ADN previamente sintetizados. Entonces alarga estas cadenas hasta rellenar la brecha donde estaba el primer.
La segunda labor consiste en unir todos los fragmentos de ADN, lo que está a cargo de la enzima ADN ligasa.
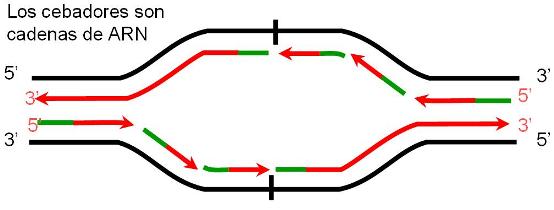 |
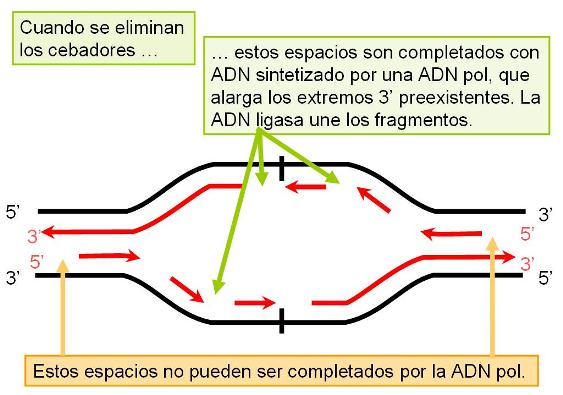 |
La autoduplicación del ADN, proceso sumamente complejo que involucra a un sinnúmero de enzimas, se cumple con una tasa muy baja de errores en cada ciclo celular. Cada 10.000.000 de nucleótidos añadidos, sólo uno lleva una base erróneamente apareada. Esto es muy importante, pues una alteración en la secuencia de bases (mutación) puede cambiar los mensajes genéticos. ¿Cómo se logra tal grado de eficiencia en la autoduplicación? Esto se debe a la actividad exonucleasa 3’->5’ de las ADN pol. Las ADN pol, cada vez que agregan un nucleótido al extremo 3’ de una cadena, realizan “una lectura de prueba”. Antes de agregar el nucleótido siguiente, revisan que el último esté correctamente apareado con el molde. De haber ocurrido un error, la enzima lo detecta y corrige, eliminando el nucleótido mal apareado en el extremo 3’ de la cadena, gracias a su actividad exonucleasa.
Telomerasa Regresar
Durante la duplicación del ADN, una enzima ADN pol sintetiza los fragmentos de ADN que van a ocupar el sitio vacío, después de la eliminación de los ARN cebadores. Esta enzima alarga los fragmentos de ADN ya existentes, agregando los nucleótidos complementarios al molde en el extremo 3’ de la cadena. Así, las cadenas nuevas se van extendiendo, de 5’ a 3’, hasta rellenar el hueco.
Sin embargo, los extremos 5’ de ambas cadenas nuevas, en los telómeros del cromosoma, no pueden ser “rellenados” por la enzima, pues allí no cuenta con ninguna cadena que le proporcione un extremo 3’ que alargar. Ese sector de la cadena hija no se sintetiza.
Por consiguiente, los dos cromosomas hijos van a tener una cadena (la sintetizada de novo) con el extremo 5’ más corto que el original.
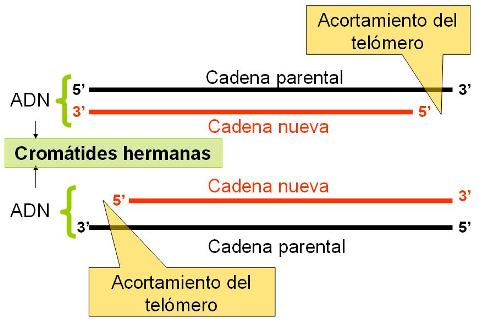 |
Este acortamiento de los telómeros se repite en el siguiente ciclo celular, en las células hijas que heredan los cromosomas. En consecuencia, con el correr de las generaciones celulares, los telómeros van sufriendo un acortamiento progresivo.
El acortamiento de los telómeros está relacionado con la senescencia o envejecimiento celular. Se ha comprobado que células en cultivos in vitro (fuera del organismo) se dividen una cierta cantidad de veces, pero luego envejecen y mueren.
La telomerasa es una enzima que alarga los telómeros, compensando la pérdida producida en cada ciclo celular. Las células que poseen telomerasa activa se comportan como líneas inmortales, esto es: se dividen indefinidamente sin envejecer. La telomerasa activa se encuentra en los organismos unicelulares, las células embrionarias y las células cancerosas.
Mitosis y Citocinesis Regresar
La mitosis es un tipo de división característico de las células eucariontes. La mitosis se inicia en una célula después de la interfase, de manera que sus cromosomas ya se encuentran duplicados. Cada cromosoma consta de dos cromátides hermanas, es decir dos copias de ADN idénticas. Durante el transcurso de la mitosis dichas copias se separan una de otra, constituyéndose, cada una de ellas, en un cromosoma hijo. Los dos grupos de cromosomas hijos están destinados a las dos células descendientes. Generalmente la mitosis va acompañada de un proceso de citocinesis o división del citoplasma. La mitosis genera dos células hijas genéticamente idénticas. En algunos casos, la célula madre forma dos núcleos hijos, pero no divide el citoplasma; esto da origen a una célula binucleada.
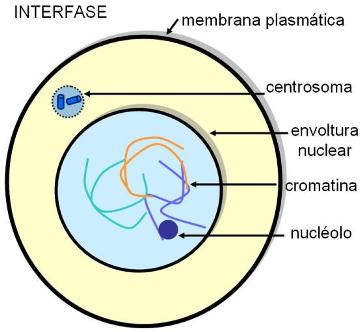 |
Durante la interfase el núcleo se encuentra organizado. El material genético adopta un estado más laxo. El nucléolo es bien visible. |
Las etapas de la mitosis son:
|
Profase: la célula adquiere una forma redondeada debido a cambios en su citoesqueleto. En esta etapa se inicia el proceso que lleva a la cromatina a su máximo grado de empaquetamiento, por lo cual los cromosomas comienzan a ser cada vez más visibles. Las asas de cromatina pertenecientes a distintos cromosomas que formaban el nucléolo se retiran y el nucléolo se dispersa.
La envoltura nuclear se disgrega en pequeñas vesículas que se incorporan al REG. La disgregación de la envoltura nuclear depende de un cambio químico disparado en la lámina nuclear, que le da sostén a la envoltura.
En el citoplasma, los centrosomas con sus centríolos previamente duplicados se empiezan a desplazar hacia polos opuestos. Algunos microtúbulos rodean a los centríolos como rayos, formando el áster. Entre ambos pares de centríolos crece el huso mitótico o acromático, a partir de la reorganización de los microtúbulos citoplasmáticos. Los centríolos, el áster y el huso conforman el aparato mitótico. En el huso mitótico hay dos tipos de fibras: las cromosómicas o cinetocóricas, que se extienden desde un centrosoma hasta los cromosomas, a los que se unirán por medio de sus cinetocoros, y las polares, que se extienden de polo a polo.
Al producirse la disgregación completa de la envoltura nuclear, los cromosomas quedan libres en el citoplasma y se inicia la siguiente etapa. |
|
|
Prometafase: las fibras del huso comienzan a invadir la zona nuclear. Las cromátides hermanas de un cromosoma se unen a fibras cromosómicas del huso provenientes de polos opuestos. Lo hacen a través de sus cinetocoros. Ocurren procesos de polimerización y despolimerización en los microtúbulos que forman las fibras del huso, lo que provoca que éstas “tironeen” de los cromosomas y los movilicen. |
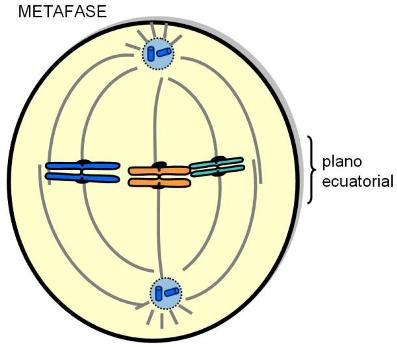 |
Metafase: los cromosomas, unidos a las fibras cromosómicas, quedan alineados en el plano medio o ecuatorial de la célula. Los cinetocoros de las cromátides hermanas en un mismo cromosoma se orientan hacia polos opuestos. De la ubicación de los cromosomas en esta etapa depende que la separación del material genético entre las futuras células se lleve a cabo correctamente. |
|
Anafase: durante las fases anteriores las cromátides hermanas permanecen unidas por un material que actúa como un pegamento, la cohesina. La cohesina se degrada en la anafase. La degradación de la cohesina permite la separación de las cromátides hermanas. Ésta se produce debido a que las fibras cromosómicas se acortan, arrastrando a los cromosomas que llevan unidos. Así, las cromátides hermanas, tironeadas por las fibras cromosómicas, inician su migración hacia polos opuestos. Cada cromátide, una vez separada de la otra, ya puede ser llamada cromosoma hijo. Los cromosomas hijos se doblan en ángulo a la altura del centrómero mientras son transportados hacia los polos. El cinetocoro, de donde traccionan las fibras cromosómicas, es la parte del cromosoma que primero avanza. Al mismo tiempo que se acortan las fibras cromosómicas, se alargan las fibras polares. Esto causa un alargamiento de la célula y un alejamiento de los polos y contribuye aun más a la separación de los cromosomas hijos. La anafase culmina cuando los cromosomas hijos llegan a los polos.
|
|
Telofase: En cada polo los cromosomas hijos son rodeados por una envoltura nuclear que se organiza a su alrededor. Los cromosomas se relajan o descondensan y empiezan a hacerse más tenues, hasta adquirir el aspecto interfásico. Se reorganiza el nucléolo en cada núcleo hijo. Se desarma el aparato mitótico, aunque persisten algunos microtúbulos en el plano ecuatorial, formando el cuerpo medio.
|
División del citoplasma |
|
Citocinesis: La citocinesis o citodiéresis es la separación del citoplasma. En las células animales, la citocinesis empieza a advertirse ya desde la anafase por la formación de un surco de segmentación en la superficie celular. En la parte media del citoplasma se origina un anillo contráctil. Se trata de una estructura formada por microfilamentos de actina en asociación con la miosina, su proteína motora. La contracción del anillo estrangula el citoplasma hasta producir la separación de las dos células hijas.
|
|
Funciones de la mitosis Regresar
En los organismos animales, la mitosis se lleva a cabo en las células somáticas o corporales. La mitosis permite:
-el crecimiento por aumento en el número de células,
-la reparación y renovación de los tejidos, y
-la formación de un organismo multicelular a partir de la cigota, durante el desarrollo embrionario.
|
|









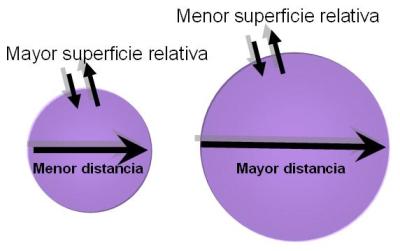
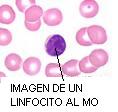
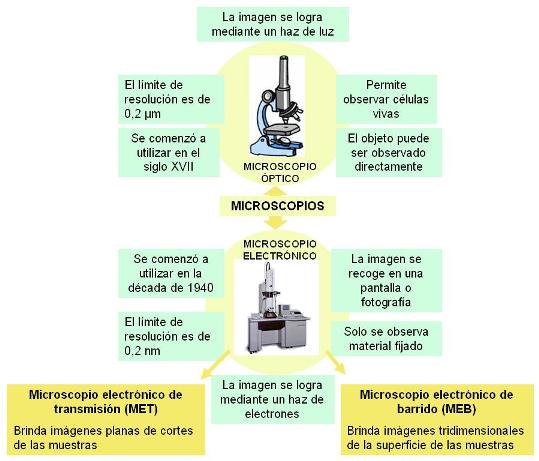

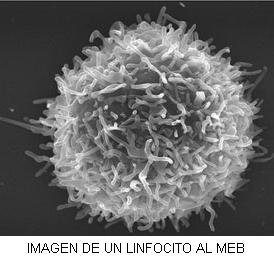
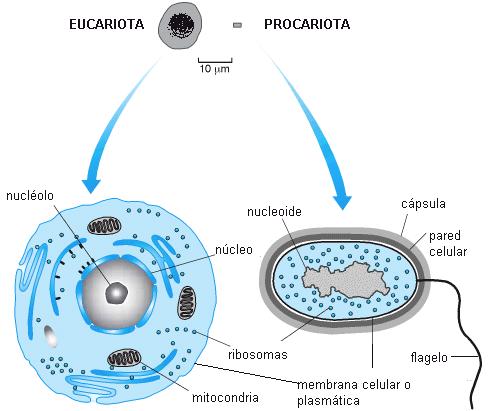
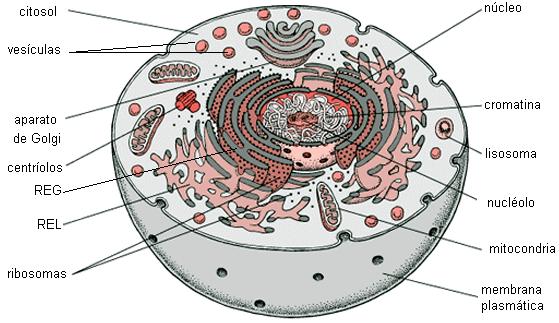
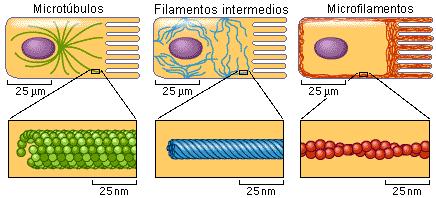
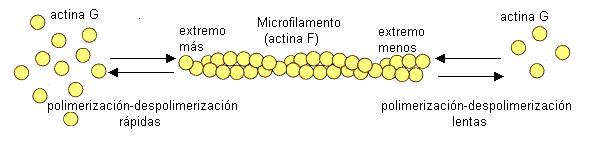
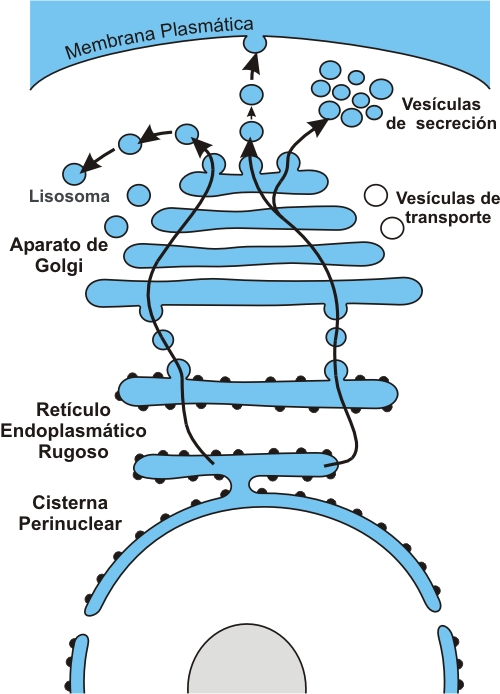
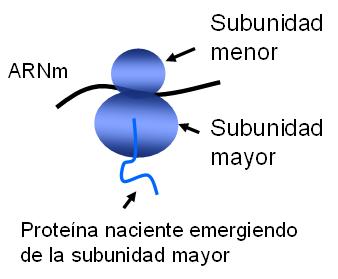
.jpg)