FOTOSÍNTESIS
Silvia Márquez - Enrique Zabala
INTRODUCCIÓN
La compleja estructura celular y su funcionamiento sólo pueden mantenerse con el aporte de materia y energía del medio externo. Sin ese aporte, las células no podrían vivir. La principal fuente de energía y materia que tienen las células, la constituyen los nutrientes orgánicos (alimento). La materia y energía contenidas en estas moléculas son el punto de partida de millares de reacciones químicas que tienen lugar dentro de la célula en un momento dado. El conjunto de estas reacciones recibe el nombre de METABOLISMO (del griego "metabole": cambio).
Prácticamente en todas las reacciones químicas que ocurren en una célula participan enzimas específicas. Estas reacciones pueden agruparse en una serie de pasos que se denominan vías. Cada vía cumple una función en la vida global de la célula. Además ciertas vías tienen muchos pasos en común.
Muchas células tienen vías que les son exclusivas, como las células vegetales que dedican gran parte de su energía a construir sus paredes celulares, actividad que no realizan las células animales. Pero, es sorprendente que en gran parte hasta el metabolismo de los más distintos organismos es muy similar. Algunas vías como la glucólisis y la respiración son prácticamente universales y existen en casi todos los sistemas vivientes. Las reacciones metabólicas pueden diferenciarse en dos tipos principales:
ANABÓLICAS: son reacciones de síntesis de moléculas relativamente complejas (por ejemplo: proteínas, polisacáridos, ácidos nucleicos) y de sus monómeros (aminoácidos, monosacáridos, nucleótidos), a partir de moléculas precursoras más sencillas.
Las reacciones anabólicas requieren el aporte de energía.
CATABÓLICAS: son reacciones de degradación de moléculas relativamente complejas (por ejemplo: monosacáridos, lípidos, etc.), procedentes del medio extracelular o de sus depósitos de reserva propios; esas moléculas son transformadas en moléculas más simples. Las reacciones catabólicas van acompañadas por la liberación de energía y proporcionan materias primas para los procesos anabólicos.
Las reacciones que requieren el aporte de energía son aquellas en las cuales los productos tienen un contenido energético mayor que los reaccionantes. Las reacciones consumidoras de energía se denominan ENDERGÓNICAS (o "cuesta arriba").
Las reacciones endergónicas típicas son las síntesis, por ejemplo:
![]()
En las células se llevan a cabo numerosas actividades que requieren un aporte energético: en ellas subyacen reacciones de tipo endergónico. Por ejemplo: la contracción muscular, el movimiento de cilias y flagelos, el transporte activo a través de membranas, el transporte en masa, la bioluminiscencia de algunas bacterias, algas y animales.
Las reacciones que liberan energía son aquellas en que la energía que poseen los productos es menor que la contenida en las sustancias reaccionantes. Las reacciones productoras de energía se denominan EXERGÓNICAS (o "cuesta abajo").
Las reacciones exergónicas son catabólicas, de degradación.
La célula usa la energía liberada en las reacciones exergónicas para impulsar las endergónicas. Esto es lo que denominamos ACOPLAMIENTO ENERGÉTICO.

Sin embargo, por lo general no se realizan acoplamientos directos entre procesos exergónicos y endergónicos. Lo más frecuente es que la célula emplee un intermediario energético. En todo tipo de células este intermediario es el ATP (adenosin-tri-fosfato). Es importante remarcar que el ATP no es un reservorio o depósito de energía, sino que actúa como un transportador de energía, desde las reacciones en que ésta se libera hacia los procesos que la consumen.
Volvamos a considerar las ecuaciones anteriores, incluyendo la intervención del ATP.
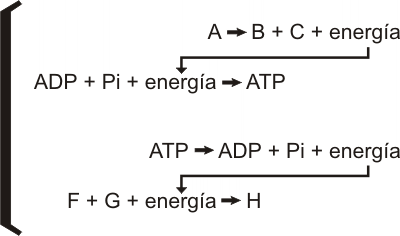
En esencia, las reacciones químicas son transformaciones en las cuales la energía almacenada en los enlaces químicos se transfiere a otros enlaces químicos recién formados. En tales transferencias, los electrones pasan de un nivel energético a otro. En muchas reacciones los electrones se transfieren de un átomo o molécula a otro. Estas reacciones muy importantes en los sistemas vivientes se conocen como reacciones de óxido-reducción (redox).
La OXIDACIÓN en los sistemas biológicos es una deshidrogenación, es decir, la pérdida de un átomo de hidrógeno (H), o lo que es equivalente la pérdida de un protón (H+) y su electrón (e-) correspondiente; o la ganancia de oxígeno. La enzima que cataliza la oxidación de una molécula debe presentar una coenzima de óxido-reducción como por ejemplo NAD+, capaz de aceptar los hidrógenos.
La REDUCCIÓN en los sistemas biológicos es una hidrogenación, es decir, la ganancia de un átomo de hidrógeno, lo que es equivalente a la ganancia de un protón (H+) más su electrón (e-) correspondiente, o la pérdida de oxígeno. La enzima que cataliza la reducción de una molécula debe presentar una coenzima reducida como por ejemplo NADH, capaz de ceder hidrógenos.
La oxidación y la reducción siempre ocurren simultáneamente porque los electrones que pierde un átomo o molécula oxidados son aceptados por otro átomo o molécula que se ha reducido en el proceso.
FOTOSÍNTESIS
Los consumidores dependen de otros seres vivos como fuente de energía. Prácticamente toda esa energía proviene de las plantas y algas. Estos productores convierten la energía solar en energía química, a través de un proceso denominado FOTOSÍNTESIS. La energía química derivada de la fotosíntesis, es almacenada en las células de esos productores en forma de hidratos de carbono y otras moléculas orgánicas necesarias para sostener todas las formas de vida del planeta.
ABSORCIÓN DE LA LUZ
A la superficie de la tierra llegan distintos tipos de radiaciones electromagnéticas. La luz es una radiación electromagnética en una banda específica de longitud de onda. La luz visible comprende una región específica del espectro electromagnético desde los 400 nm. a 700 nm. de longitud de onda. Dentro de este espectro, el violeta tiene la longitud de onda más corta, mientras que la luz roja tiene la más larga.
La luz está formada por partículas de energía llamada FOTONES. La energía de un fotón es distinta para la luz de las distintas longitudes de onda. Cuanto más corta es dicha longitud, mayor es la energía de la luz. Por el contrario, en las longitudes de onda largas hay menos energía. Resumiendo, la energía del fotón es inversamente proporcional a la longitud de onda. La clorofila refleja la luz de longitud de onda comprendida entre los 500 y 600 nm. (a ello se debe su color) y absorbe de una manera máxima las ondas de color azul violáceo y rojo. Estas ondas son las que producen la mayor actividad fotosintética.
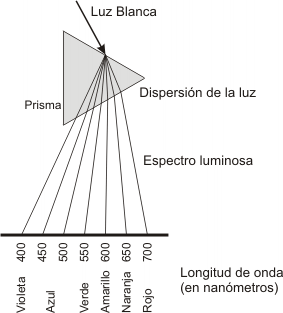
Fig. 8.1 - Dispersión de la luz visible. El prisma separa el rayo en bandas de diferentes colores que van del rojo, en un extremo, al violeta , en otro. Cada color corresponde a una longitud de onda diferente. Dentro del espectro visible, las ondas de mayor longitud de onda corresponden al rojo y son las que se desvían menos al pasar a través del prisma. Las ondas de longitud más cortas pertenecen al color violeta y son las que más se desvían.
La luz puede excitar ciertos tipos de moléculas, y por lo tanto, desplazar electrones hacia niveles de energía superiores. Cuando un electrón pasa a un nivel de energía más elevado, se dice que el átomo está excitado. Esta excitación puede ser el resultado de la absorción de cualquier tipo de energía. En la fotosíntesis, por supuesto, la energía que provoca la excitación proviene del sol.
Si un fotón tiene suficiente energía para producir esta excitación, pueden ocurrir dos cosas:
1) El electrón regresa pronto a su nivel original. La energía se disipa generalmente como calor o luz de longitud de onda más larga.
2) El electrón se pierde dejando al átomo con una carga positiva neta. El electrón emitido puede ser aceptado por un agente reductor, lo que deja al átomo excitado, con una carga positiva neta.
LOS PIGMENTOS CAPTADORES DE ENERGÍA
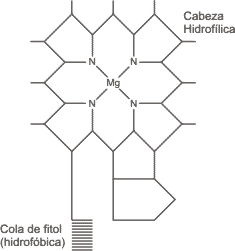
Fig. 8.2 - Esquema de la estructura química de la clorofila
La clorofila es el pigmento que les da el color verde a las plantas; aunque también está presente en plantas y algas de distintos colores que hacen fotosíntesis. La clorofila se excita con la luz o con la energía que pasa desde otros pigmentos excitados por la luz, a los que se llama pigmentos accesorios.
Existen varios tipos de clorofila. Es factible que el más importante sea la clorofila a. Esta molécula presenta una cabeza hidrofílica que es un anillo de porfirina, que tiene un átomo central de Mg (Magnesio), y una cola o cadena lateral formada por un fitol (terpeno lineal), que es hidrofóbica, lo que determina la orientación de la molécula de clorofila en las membranas internas del cloroplasto.
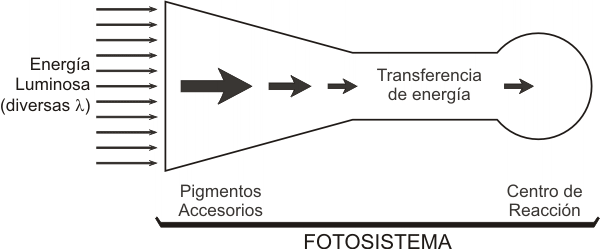
Todos los organismos fotosintetizadores contienen un pigmento orgá-nico capaz de absorber la radiación visible e iniciar las reacciones fotoquímicas de la fotosíntesis. Este pigmento es la clorofila a.
En colaboración con ella actúan otros tipos de clorofila y diversos pigmentos, que comparten una propiedad esencial: cambian su configuración electrónica cuando reciben luz de cierta longitud de onda. Estos pigmentos accesorios absorben energía radiante y la canalizan hacia la clorofila a. La siguiente es una lista de los principales pigmentos fotosintéticos y los organismos donde se encuentran.
|
Cuadro 8.1 - Pigmentos fotosintÉticos
|
|
|
Tipo de pigmento |
Organismos donde se encuentra |
|
Clorofilas clorofila a clorofila b clorofila c clorofila d |
· Todas las plantas superiores y todas las algas · Todas las plantas superiores y las algas verdes · Diatomeas y algas pardas · Algas rojas |
|
Carotenoides b-caroteno a-caroteno luteína xantófila ficoxantina |
· Plantas superiores y la mayoría de las algas · La mayoría de las plantas y algunas algas · Algas verdes, algas rojas y plantas superiores · Plantas superiores · Diatomeas y algas pardas |
|
Ficobilinas ficoeritrinas ficocoaninas |
· Algas rojas y algunas algas azul-verdes · Algas azul-verdes y algunas algas rojas |
Todos estos pigmentos que acompañan a la clorofila a, son receptores suplementarios de luz que encauzan la energía que han absorbido. La transferencia de electrones se hace de pigmento a pigmento, hasta llegar a uno cuya máxima absorción se realiza a la longitud de onda más alta y llega a la clorofila a formando un fotosistema.
ESTRUCTURA DEL CLOROPLASTO
Los cloroplastos son organoides en los que se lleva a cabo la fotosíntesis en las células vegetales eucariontes. En los procariontes fotosintetizadores no existen tales organoides. Los pigmentos captadores de energía, se hallan asociados a las laminillas derivadas de la membrana plasmática.
Generalmente poseen forma discoide, su tamaño es variable (término medio: 4 a 6 nm de longitud). El cloroplasto está limitado por una doble membrana que no posee clorofila ni citocromos, que envuelve a la matriz o estroma. Esta posee el 50% de las proteínas, que en su mayoría son solubles; también contiene ribosomas y ADN de características procarionte. Estas dos últimas características explican que el cloroplasto sea un organoide semiautónomo.
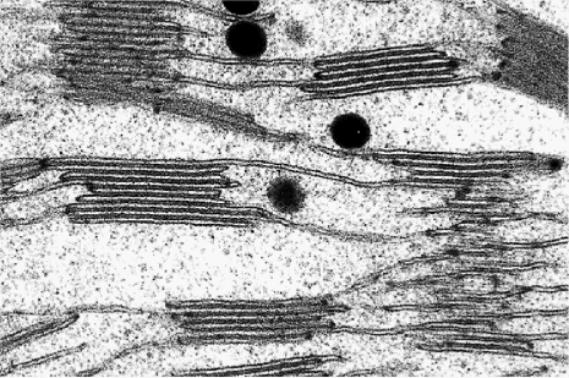
Dentro del estroma se hallan los tilacoides, que son vesículas aplanadas dispuestas como un retículo membranoso. Su superficie externa está en contacto con el estroma, la interna limita el espacio intratilacoide. Los tilacoides se disponen como pilas de monedas para formar las granas, entre las cuales se extienden laminillas intergrana formando un retículo membranoso.
La membrana tilacoidal responde al modelo del mosaico lipoproteico. Se encuentra en ella el 50% de los lípidos del cloroplasto y entremezclados moléculas de clorofila, carotenoides, plastoquinonas que intervienen en la fotosíntesis.
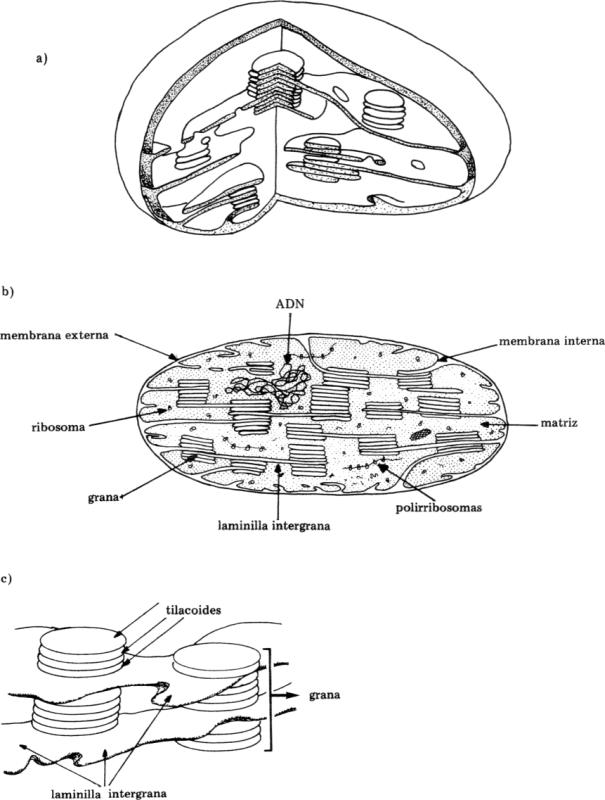
Fig. 8.4 - Ultraestructura del cloroplasto. (a) esquema tridimensional, (b) esquema de la ultraestructura al M.E.T, (c) grana de un cloroplasto
Las moléculas de clorofila se disponen agrupadas con proteínas intrínsecas formando varios complejos de gran importancia en la fotosíntesis (fotosistemas I y II). Además de estos complejos, hay otros complejos moleculares que no poseen- clorofila, a saber:
1) Complejo cit f b6 (contiene cit f y b6 y plastocianina, proteína que posee Cu2+). Este complejo interviene en la cadena de electrones.
2) Complejo CF0 o HF0, (contiene proteínas hidrofóbicas). Actúa en la traslocación de protones y se une a CF1 para formar el complejo ATPasa.
Las partículas CF1, son factores de acoplamiento, que intervienen en la fotofosforilación de ADP a ATP. Están localizadas en la superficie externa de la membrana tilacoidal. Se hallan unidas a la membrana por medio del segmento CF0, que representa un canal protónico formado por proteínas hidrofóbicas.
ETAPAS DE LA FOTOSÍNTESIS
a) Etapa lumÍnica o dependiente de la luz
Consiste en la transferencia de energía lumínica en química bajo la forma de ATP, y en la obtención de una fuente reductora de alta energía: la coenzima NADPH.
Como subproducto de esta etapa se obtiene 02 . La etapa lumínica se lleva a cabo en las granas.
La etapa lumínica, se desencadena cuando el fotosistema I (PS I) absorbe un fotón, este PS I emite un electrón que es aceptado por una proteína, la Ferredoxina. Este fotosistema queda por lo tanto con carga positiva. La ferredoxina ahora reducida, transporta electrones al NADP+ el cual, juntamente con H+ provenientes de la fotooxidación del H2O, es reducido a NADPH.
Por otro lado, el PS II también es excitado por la luz y sus electrones son llevados a un nivel de alta energía donde son aceptados sucesivamente por una cadena transportadora específica. Finalmente los electrones son aceptados por el PS I que había quedado positivo, restituyendo así su estado inicial.
El flujo de electrones desde el PS II al I es un transporte exergónico, esta energía se emplea para bombear H+ a través de la membrana tilacoidal hacia el interior de ésta. Los H+ regresan luego desde el interior hacia el estroma a través de canales especiales de la membrana formados por los complejos CF0 Y CF1.
Mientras tanto el PS II que había quedado con carga positiva, para recuperar su estado inicial, promueve la oxidación (fotólisis) del H2O y capta sus electrones, quedando así restituida su carga eléctrica. Pero además se producen H+ que como ya se mencionara anteriormente, contribuyen ala reducción de la coenzima NADPH. El O2 que resulta de la oxidación de la molécula de H2O, se libera al medio.
Reacción de Hill
Una prueba de que el oxígeno liberado en la fotosíntesis proviene del agua y no del dióxido de carbono se debe a los experimentos realizados por Robín Hill (1937), quien obtuvo oxígeno al exponer cloroplastos aislados a la luz en presencia de un aceptor de electrones, el ferricianuro, pero en ausencia de CO2 y NADP. Esto demostró que el oxígeno también se libera sin que haya reducción del CO2, cuando se dispone de un aceptor de electrones. La ausencia de CO2 en la mezcla confirmó que el oxígeno liberado proviene de las moléculas de agua. Además, los experimentos demostraron que el fenómeno fundamental en la fotosíntesis es la transferencia inducida por la luz, de electrones de un compuesto a otro.
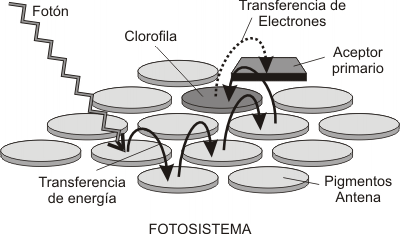
Fig. 8.6 - Transferencia de la energía de excitación. La energía de excitación se transfiere al azar a pigmentos accesorios (antena) que absorben luz de longitud de onda creciente hasta alcanzar el centro de reacción en la clorofila, que transfiere un electrón excitado a un aceptor primario.
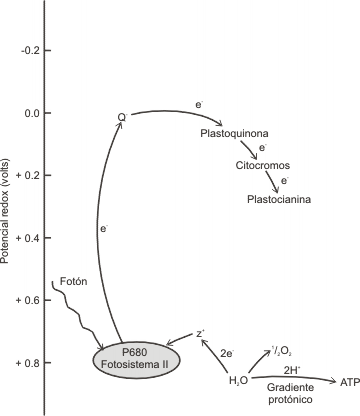
Fig. 8.7 - Flujo de electrones en el fotosistema II. Cuando el PSII es activado por la absorción de un fotón, se produce un poderoso oxidante, llamado Z+. El potencial de este oxidante es capaz de extraer electrones del agua, formando O2 y contribuyendo a la formación de un gradiente protónico que genera ATP. Los electrones fluyen luego hacia el reductor Q-, que está fuertemente unido a las membranas tilacoidales. Luego el electrón pasa hacia la plastoquinona soluble y de ahí a los otros aceptores
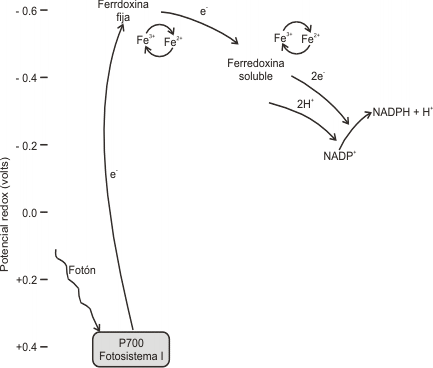
Fig. 8.8 - Paso de los electrones desde el fotosistema I hacia el NADP+.
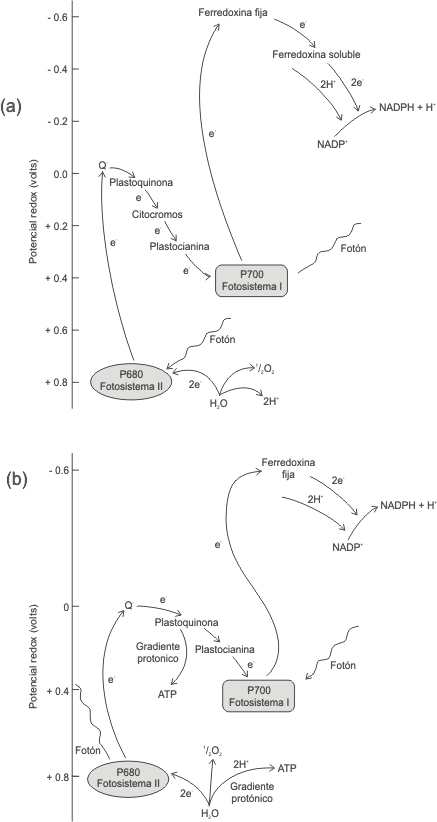
Fig. 8.9 - (a) Esquema en el que se muestra la relación que hay entre el fotosistema I y II, a través de la cual se recuperan los electrones transferidos al NADPH en el PSI, ya que pasan a este los electrones derivados del H2O en el PSII. (b) Flujo de electrones desde el H2O hacia el NADP+, para la formación del NADPH.
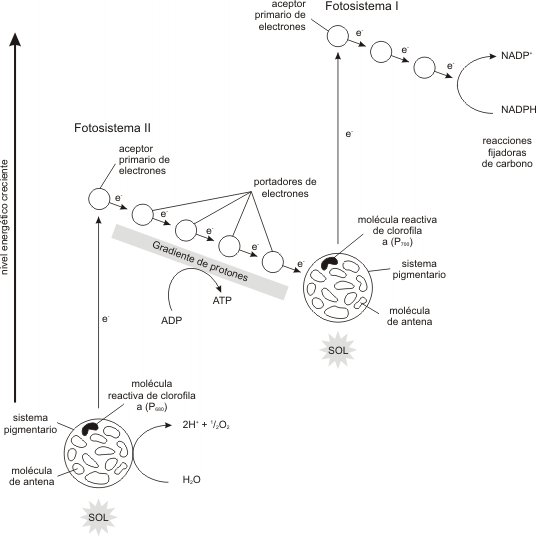
Fig. 8.10 - Esquema simplificado de la etapa lumínica de la fotosíntesis
|
La energía luminosa de la fotosíntesis consiste en la conversión de: ENERGÍA
LUMINOSA La energía química queda contenida en moléculas de dos tipos: [ ATP [ NADPH Además. Como subproducto de esta etapa , se obtiene: [ O2 (oxígeno molecular) |
FORMACIÓN DE ATP EN LOS GRADIENTES PROTÓNICOS
QUIMIÓSMOSIS
Conforme a la teoría quimiosmótica, los organoides que usan energía (cloroplastos), acumulan protones dentro de compartimentos membranosos especiales. Los protones difunden luego a través de las membranas (como ya se mencionara en la etapa lumínica). La energía potencial de la membrana creada por las diferencias en la concen-tración de protones es captada por el complejo proteico CF0-CF1. Este complejo usa una parte de la energía para sintetizar ATP.
Según pasan los electrones a través de la cadena de aceptores los protones son bombeados hacia el interior del tilacoide donde se acumulan. Los protones equivalen a iones de H+, por lo tanto el pH interior del tilacoide desciende, con lo que se establece una diferencia de pH a través de la membrana.
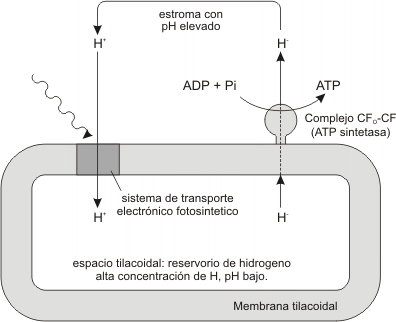
Fig. 11- Gradiente de protones a través de la membrana del tilacoide
Los iones hidrógeno altamente concentrados en el interior del tilacoide tienden a difundirse hacia el exterior, pero no lo pueden hacer porque la membrana es impermeable a ellos, excepto a través del complejo CF0 – CF1. Este complejo actúa como una especie de conducto a través de los cuales los protones pueden salir. A medida que los protones pasan a través del complejo van liberando su energía, la cual se utiliza para la síntesis de ATP.
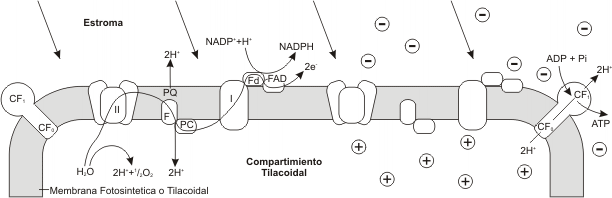
Fig. 8.12 - Modelo tentativo de la organización de la membrana tilacoidal. Los PSI y PSII absorben luz de longitudes de onda distintas. En el PSII se disocia el agua en dos protone (H+), oxígeno y dos electrones (e-). Los electrones se transfieren a través de una cadena de transporte de electrones que consta de plastoquinona, plastocianina, el PSI, ferredoxina y Flavin-adenin-dinucleótido, hasta que, con la ayuda de un protón, reducen el NADP+ a NADPH en la superficie externa de la membrana del tilacoide. A la derecha del esquema, observamos una acumulación de cargas positivas en la zona interna, que proceden de dos fuentes: los protones liberados por la disociación del agua y cargas positivas que parecen ser translocadas desde el exterior a través de la membrana durante el transporte de electrones. El flujo de protones hacia el exterior a través de los factores de acoplamiento (CF0 y CF1) permite la síntesis de ATP a partir de ADP.
b) Etapa Oscura o Ciclo de Calvin (C3)
Consiste en la reducción de moléculas de CO2 para formar glúcidos mediante las fuentes de energía (ATP) y la fuente reductora (NADPH) obtenidas en la etapa clara. La etapa oscura ocurre en la matriz del cloroplasto con la intervención de numerosas enzimas que actúan en un ciclo.
En el estroma existe una molécula que acepta la adición de CO2: la ribulosa difosfato (de 5 carbonos) que forma un compuesto transitorio de 6 C, que rápidamente de hidroliza dando 2 moléculas de fosfoglicerato (PGA) (cada molécula de PGA contiene 3 átomos de carbono y de ahí el nombre de vía de 3 carbonos). La enzima que cataliza esta reacción es la ribulosa difosfato carboxilasa (RUBISCO) localizada en la superficie estromal de las membranas tilacoidales.
El PGA se convierte en difosfoglicerato mediante el gasto de un ATP, es necesaria la participación del NADPH para reducir al difosfoglicerato y convertirlo en gliceraldehído fosfato (PGAL), un azúcar de 3 C. Dos de estas triosas (PGAL) se condensan y forman una hexosa: la fructosa 1,6 difosfato, a la brevedad uno de los grupos fosfatos es eliminado enzimáticamente para producir fructosa 6 fosfato, la cual experimenta un reacondicionamiento molecular para convertirse en glucosa 6 fosfato. Este último compuesto puede ser incorporado a una molécula de almidón para ser almacenado.
Para que continúe el ciclo es necesario regenerar la ribulosa difosfato. El resto de las moléculas de PGAL se destinan a la regeneración de este compuesto. Durante esta etapa se producen condensaciones, hidrólisis y reordenamientos, con intermediarios de 3, 4, 5, 6, 7 carbonos. Finalmente se forma ribulosa monofosfato que mediante gasto de ATP, es fosforilada a ribulosa difosfato, con lo cual se cierra el ciclo.
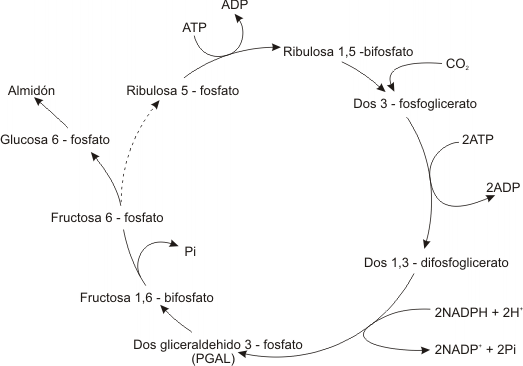
Para sintetizar una molécula de glucosa se necesitan 6 vueltas del ciclo de Calvin puesto que en cada una de ellas se reduce una molécula de CO2. Para fosforilar 12 moléculas de PGAL y convertirlas en 12 moléculas de difosfoglicerato se necesitan 12 ATP, mientras que se emplean 12 de NADPH para reducir 12 moléculas de difosfoglicerato a gliceraldehído fosfato. Luego se consumen otras 6 moléculas de ATP en la regeneración de la ribulosa difosfato.
Ciclo de C4 de Hatch y Slack
Otra de las vías alternativas para la incorporación del CO2 se realiza en un grupo numeroso e importante de plantas entre ellas maíz, sorgo, caña de azúcar. Estas plantas tienen una alta eficiencia en la captación de CO2 y requieren relativamente menos agua.
La vía del C4 ocurre en cloroplastos de morfología diferente (se caracterizan por poseer una extensa red de tilacoides organizados en grana bien desarrolladas).
En ellos el CO2 de la atmósfera es incorporado a una molécula de 3C, el fosfoenolpiruvato (PEP) que se convierte en un compuesto de 4C, el ácido oxalacético. Esta reacción es catalizada por la enzima PEP carboxilasa. Luego el ácido oxalacético se reduce a ácido málico o, mediante la adición de un grupo amino, se convierte en ácido aspártico. Este compuesto es transportado a un cloroplasto típico. Allí pierde CO2 (se descarboxila) para producir CO2 y ácido pirúvico. Entonces el CO2 entra en el ciclo de Calvin.
Cabe preguntarse por qué este tipo de plantas emplean este mecanismo energéticamente tan costoso y complicado. El CO2 no está disponible en todo momento porque éste entra en la hoja por los estomas que son poros que se abren y cierran de acuerdo a las necesidades del vegetal. Los estomas tienen que permanecer cerrados gran parte del tiempo para evitar que la planta pierda agua por transpiración.
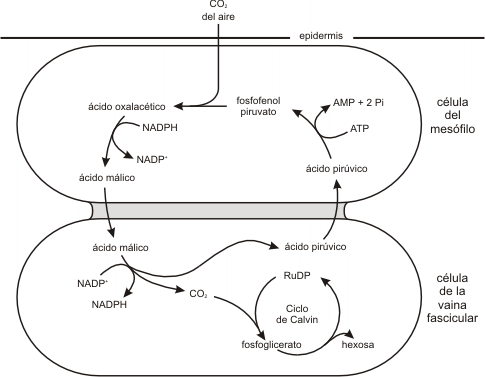
Fig. 8.14 - Vía de fijación de carbono en las plantas C4. Primero se fija CO2 como ácido oxalacético en las células del mesófilo. Luego este CO2 se transfiere a las células de la vaina fascicular, donde libera CO2. Este último entra en el ciclo de Calvin. El ácido pirúvico retorna a la célula del mesófilo, donde se fosforila a PEP.
La PEP caboxilasa posee mayor afinidad por el CO2 que las RUDP carboxilasa o Rubisco (enzima que cataliza la incorporación del CO2 a la ribulosa di P). Incluso a bajas concentraciones de CO2, la enzima funciona rápidamente para fijarlo al PEP. En comparación con la Rubisco, fija el CO2, más pronto y a niveles más bajos, manteniendo más baja su concentración dentro de la hoja. Esto provoca que se cree un gradiente de concentración entre las células y su medio ambiente. Por lo tanto cuando la planta abre sus estomas el CO2 difunde rápidamente con gran eficiencia al interior de la hoja. En consecuencia las plantas C4 poseen una gran venta en las regiones cálidas y áridas.
Resumiendo, la función del C4 es aumentar la cantidad de CO2 incorporado a la atmósfera, en condiciones bajo las cuales no puede intervenir eficazmente la ribulosa difosfato (C3). La síntesis de glúcidos a partir del CO2 fijado vía C3, o vía C4, se realiza a través del ciclo de Calvin.
|
Cuadro 8.2 - Resumen de las ecuaciones de la fotosÍntesis
|
|
Ecuación general de las reacciones dependientes de la luz: 12
H2O + 12 NADP+ + 18 ADP + 18 Pi Ecuación general de las reacciones independientes de la luz: 12
NADPH + 12 H+ + 18 ATP + 6 CO2 Procediendo a la simplificación de los elementos comunes en ambos lados de las ecuaciones acopladas, se obtiene la ecuación global simplificada de la fotosíntesis: 6 CO2
+ 12 H2O
|
|
Cuadro 8.3 - Resumen de las principales reacciones de la fotosíntesis
|
|||
|
Serie de reacciones |
Resumen del proceso |
Materiales necesarios |
Productos finales |
|
Reacciones dependientes de la luz (en la membrana tilacoidal) |
Se utiliza la energía de la luz solar para dividir el agua, sintetizar ATP y reducir el NADP |
||
|
Reacciones fotoquímicas |
Se energiza la clorofila, el centro de reacción envía un electrón energizado hacia un aceptor de electrones
|
Energía lumínica, pigmentos como la clorofila |
|
|
Transporte de electrones |
Los electrones son transportados a lo largo de una cadena de aceptores de electrones en las membranas tilacoidales; los electrones , reducen el NADP+, la división del agua genera H+, que se acumulan dentro de los tilacoides.
|
Electrones, NADP+, H2. |
NADPH + H+ + O2; H+ |
|
Quimiósmosis |
Se bombea H+ a través de la membrana tilacoidad, formando un gradiente de protones; esos protones regresan a través de los conductos especiales de la membrana formados por el complejo proteínico CF0 - CF1; se produce ATP.
|
Un gradiente protónico y un potencial de membrana; ADP + Pi (fosfato inorgánico) |
ATP |
|
Reacciones independientes de la luz (en el estroma) |
Fijación de CO2. El CO2 se combina con un compuesto orgánico |
Ribulosa bifosfato, CO2, ATP, NADPH + H+
|
|
ACTIVIDADES DE EVALUACIÓN
1) En términos de la economía de la célula:
a) ¿Qué proveen a la célula los procesos anabólicos?
b) ¿Qué proveen a la célula los procesos catabólicos?
c) ¿Cómo dependen los unos de los otros?
Elabore su respuesta en forma de gráfico.
2) Complete el siguiente cuadro.
|
Ubicación celular |
Sustrato |
Producto |
Exergónica o endergónica |
|
|
Etapa clara de la fotosíntesis |
||||
|
Etapa oscura de la fotosíntesis |
3) Dibuje un cloroplasto y rotule todas sus membranas y compartimentos.
4) ¿Cuál es la función de los pigmentos accesorios?
5) De los procesos indicados en el siguiente cuadro, identifique el carácter exergónico o endergónico.
|
Proceso |
Exergónico |
Endergónico |
|
Síntesis de glucosa |
||
|
Hidrólisis de proteínas |
||
|
Síntesis de lípidos |
||
|
Transporte activo |
||
|
Endocitosis |
6) ¿Mediante qué reacción se produce O2 molecular a partir de H2O?
7) ¿Cómo se establece un gradiente protónico a través de la membrana tilacoidal? ¿Cómo se traduce esto en la síntesis de ATP?
8) ¿Cómo se producen y utilizan el ATP y el NADPH durante la fotosíntesis?
9) Justifique la veracidad o falsedad del siguiente enunciado: “Durante la fase clara se libera O2 a partir del CO2por medio de reacciones químicas que requieren ATP”.
10) ¿De qué manera la fotosíntesis C4 es ventajosa para las plantas que la realizan?
Preguntas de opción multiple
1- La incidencia de luz sobre la clorofila provoca que ésta:
a- se hidrolice liberando su cola de fitol
b- libere electrones
c- libere oxígeno al medio
d- capte electrones
e- dióxido de carbono del medio
2 - ¿Cuál de las siguientes parejas: proceso/localización no se corresponde:
a- reacción lumínica / grana
b- cadena de transporte de electrones /membrana tilacoide
c- ciclo de Calvin / estroma del cloroplasto
d- doble membrana del cloroplasto /ATP sintetasa
3- La clorofila II conocida como clorofila P680 es reducida por los electrones provenientes del:
a- Fotosistema I
b- Fotosistema II
c- Agua
d- NADPH
e- NADP
4- Los electrones que circulan a través de los dos fotosistemas tiene su menor energía potencial a nivel del:
a- P700
b- P680
c- NADPH
d- Agua
5- ¿Cuántas vueltas del ciclo de Calvin son necesarias para producir una molécula de glucosa?
a- 1
b- 2
c- 3
d- 5
e- 6
BIBLIOGRAFIA
l Alberts, B et al; (1996) Biología Molecular de la Célula. 3ra Edición. Ediciones Omega S.A. Barcelona.
l Campbell, N; (1997) Biology. 4th Edition. the Benjamin Cummings Publishing Company. Inc. California
l Castro, Handel y Rivolta . Actualizaciones en Biología. (1986). Ed. EUDEBA
l Castro R. et al. Investigación y Ciencia., N° 39, Diciembre de 1979.
l Curtis y Barnes (1992). Biología. 5ª Ed. Bs.As. Editorial Médica Panamericana.
l CONICET-SENOC (1984) Módulo 3 y 4.
l De Robertis(h); Hib; Ponzio. (1996).Biología Celular y Molecular de De Robertis. 12º Edición. El Ateneo. Bs.As.
l De Robertis, E.; Hib, J.; (1998) .Fundamentos de Biología Celular y Molecular. El Ateneo. Bs.As.
l Karp, G.; (1998) Biología Celular y Molecular. Ed. Mc Graw Hill Interamericana. México.
l Smith and Wood; (1997) .Biología Celular. Ed.Addison-Wesley, Iberoamericana S.A.
l
Solomon
y col. (1998) . Biología de Villee. 4ª. Ed. Mex. McGraw-Hill. Interamericana