RESPIRACIÓN CELULAR
Silvia Márquez - Enrique Zabala
El proceso por el cual las células degradan las moléculas de alimento para obtener energía recibe el nombre de RESPIRACIÓN CELULAR.
La respiración celular es una reacción exergónica, donde parte de la energía contenida en las moléculas de alimento es utilizada por la célula para sintetizar ATP. Decimos parte de la energía porque no toda es utilizada, sino que una parte se pierde.
Aproximadamente el 40% de la energía libre emitida por la oxidación de la glucosa se conserva en forma de ATP. Cerca del 75% de la energía de la nafta se pierde como calor de un auto; solo el 25% se convierte en formas útiles de energía. La célula es mucho más eficiente.
La respiración celular es una combustión biológica y puede compararse con la combustión de carbón, bencina, leña. En ambos casos moléculas ricas en energía son degradadas a moléculas más sencillas con la consiguiente liberación de energía.
Tanto la respiración como la combustión son reacciones exergónicas.
Sin embargo existen importantes diferencias entre ambos procesos. En primer lugar la combustión es un fenómeno incontrolado en el que todos los enlaces químicos se rompen al mismo tiempo y liberan la energía en forma súbita; por el contrarío la respiración es la degradación del alimento con la liberación paulatina de energía. Este control está ejercido por enzimas específicas.
En segundo lugar la combustión produce calor y algo de luz. Este proceso transforma energía química en calórica y luminosa. En cambio la energía liberada durante la respiración es utilizada fundamentalmente para la formación de nuevos enlaces químicos (ATP).
La respiración celular puede ser considerada como una serie de reacciones de óxido-reducción en las cuales las moléculas combustibles son paulatinamente oxidadas y degradadas liberando energía. Los protones perdidos por el alimento son captados por coenzímas.
La respiración ocurre en distintas estructuras celulares. La primera de ellas es la glucólisis que ocurre en el citoplasma. La segunda etapa dependerá de la presencia o ausencia de O2 en el medio, determinando en el primer caso la respiración aeróbica (ocurre en las mitocondrias), y en el segundo caso la respiración anaeróbica o fermentación (ocurre en el citoplasma).
GLUCÓLISIS
La glucólisis, lisis o escisión de la glucosa, tiene lugar en una serie de nueve reacciones, cada una catalizada por una enzima específica, hasta formar dos moléculas de ácido pirúvico, con la producción concomitante de ATP. La ganancia neta es de dos moléculas de ATP, y dos de NADH por cada molécula de glucosa.
Las reacciones de la glucólisis se realizan en el citoplasma, como ya adelantáramos y pueden darse en condiciones anaerobias; es decir en ausencia de oxígeno.
Los primeros cuatro pasos de la glucólisis sirven para fosforilar (incorporar fosfatos) a la glucosa y convertirla en dos moléculas del compuesto de 3 carbonos gliceraldehído fosfato (PGAL). En estas reacciones se invierten dos moléculas de ATP a fin de activar la molécula de glucosa y prepararla para su ruptura.
Paso 1
La serie de reacciones glucolíticas se inicia con la activación de la glucosa
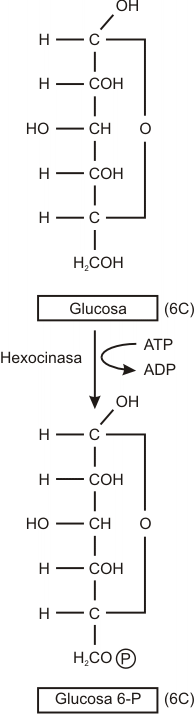
Glucosa
+ ATP ![]() glucosa
6 fosfato + ADP
glucosa
6 fosfato + ADP
La reacción del ATP con la glucosa para producir glucosa 6-fosfatoy ADP es exergónica. Parte de la energía liberada se conserva en el enlace que une al fosfato con la molécula de glucosa que entonces se energiza.
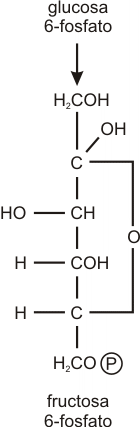
Paso 2
La glucosa 6-fosfato sufre una reacción de reordenamiento catalizada por una isomerasa, con lo que se forma fructosa 6-fosfato.
Paso 3
La fructosa 6-fosfato acepta un segundo fosfato del ATP, con lo que se genera fructosa 1,6-difosfato; es decir fructosa con fosfatos en las posicio-nes 1 y 6.
La enzima que regula esta reacción es la fosfofructocinasa.
Nótese que hasta ahora se han invertido dos moléculas de ATP y no se ha recuperado energía.
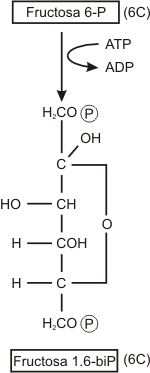
La fosfofructocinasa es una enzima alostérica, el ATP es un efector alostérico que la inhibe. La interacción alostérica entre ellos es el principal mecanismo regulador de la glucólisis. Si existe ATP en cantidades suficientes para otros fines de la célula, el ATP inhibe la actividad de la enzima y así cesa la producción de ATP y se conserva glucosa. Al agotar la célula la provisión de ATP, la enzima se desinhibe y se reanuda la degradación de la glucosa. Este es uno de los puntos principales del control de la producción de ATP.
Paso 4
La fructosa 1,6 -difosfato se divide luego en dos azúcares de 3 carbonos, gliceraldehído 3-fosfato y dihidroxiacetona fosfato. La dihidroxiacetona fosfato es convertida enzimáticamente (isomerasa) en gliceraldehído fósfato. Todos los pasos siguientes deben contarse dos veces para tener en cuenta el destino de una molécula de glucosa.
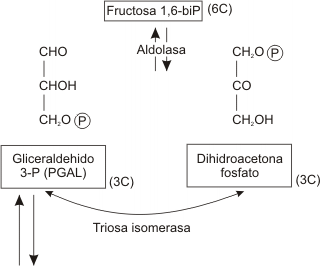
Debemos recordar que hasta el momento no se ha obtenido ninguna energía biológicamente útil. En reacciones subsecuentes, la célula recupera parte de la energía contenida en el PGAL.
Paso 5
Las moléculas de PGAL se oxidan es decir, se eliminan átomos de hidrógeno con sus electrones, y el NAD+ se reduce a NADH. Esta es la primera reacción de la cual la célula cosecha energía. El producto de esta reacción es el fosfoglicerato. Este compuesto reacciona con un fosfato inorgánico (Pi) para formar 1,3 difosfoglicerato. El grupo fosfato recién incorporado se encuentra unido por medio de un enlace de alta energía.
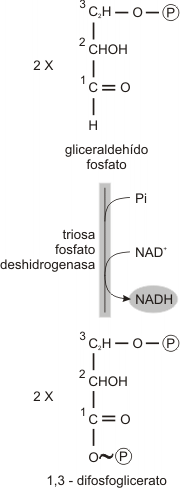
Paso 6
El fosfato rico en energía reacciona con el ADP para formar ATP. (en total dos moléculas de ATP por molécula de glucosa). Esa transferencia de energía desde un compuesto con un fosfato, de alta energía se conoce como fosforfiación.
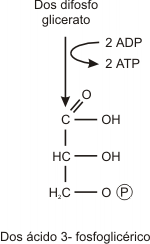
Paso 7
El grupo fosfato remanente se transfiere enzimáticamente de la posición 3 a la posición 2 (ácido 2-fosfoglicérico).
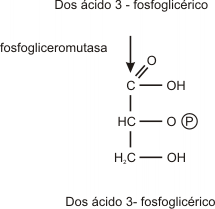
Paso 8
En este paso se elimina una molécula de agua del compuesto 3 carbono. Este reordenamiento interno de la molécula concentra energía en la vecindad del grupo fosfato. El producto es el ácido fosfoenolpirúvico (PEP).
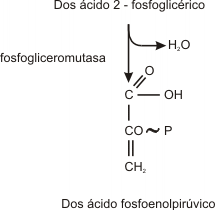
Paso 9
El ácido fosfoenolpirúvico tiene la capacidad de transferir su grupo fosfato a una molécula de ADP para formar ATP y ácido pirúvico. (dos moléculas de ATP y ácido pirúvico por cada molécula de glucosa).
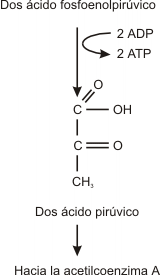
RESUMEN DE LA GLUCÓLISIS
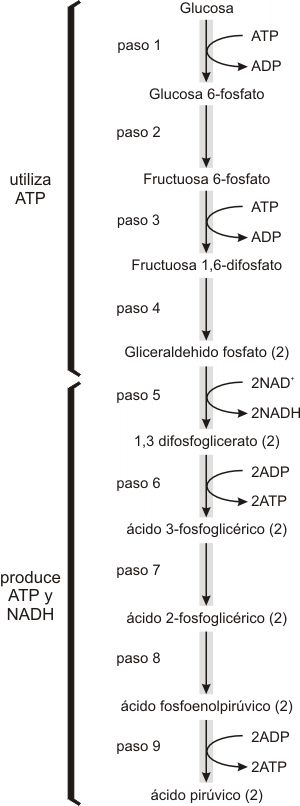
Fig. 9.1 - Resumen de las dos etapas de la glucólisis. En la primera etapa se utilizan 2 ATP y la segunda produce 4 ATP y 2 NADH. Otros azúcares, además de la glucosa, como la manosa, galactosa y las pentosas, así como el glucógeno y el almidón, pueden ingresar en la glucólisis una vez convertidos en glucosa 6-fosfato.
|
Ecuación de la Glucólisis Glucosa
+ 2 ADP + 2 Pi + 2 NAD+
|
VÍAS ANAERÓBICAS
El ácido pirúvico puede tomar por una de varias vías. Dos son anaeróbicas (sin oxígeno) y se denomina FERMENTACIÓN ALCOHÓLICA y FERMENTACIÓN LÁCTICA.
A la falta de oxígeno, el ácido pirúvico puede convertirse en etanol (alcohol etílico) o ácido láctico según el tipo de célula. Por ejemplo, las células de las levaduras pueden crecer con oxígeno o sin él. Al extraer jugos azucarados de las uvas y al almacenarlos en forma anaerobia, las células de las levaduras convierten el jugo de la fruta en vino al convertir la glucosa en etanol. Cuando el azúcar se agota las levaduras dejan de fermentar y en este punto la concentración de alcohol está entre un 12 y un 17 % según sea la variedad de la uva y la época en que fue cosechada.
La formación de alcohol a partir del azúcar se llama fermentación.
Fermentación alcohólica
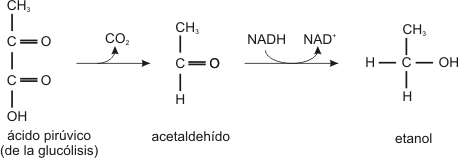
El ácido pirúvico formado en la glucólisis se convierte anaeróbicamente en etanol. En el primer caso se libera dióxido de carbono, y en el segundo se oxida el NADH y se reduce a acetaldehído.
Otras células, como por ejemplo los glóbulos rojos, las células musculares y algunos microorganismos transforman el ácido Pirúvico en ácido láctico.
En el caso de las células musculares, la fermentación láctica, se produce como resultado de ejercicios extenuantes durante los cuales el aporte de oxígeno no alcanza a cubrir las necesidades del metabolismo celular. La acumulación del ácido láctico en estas células produce la sensación de cansancio muscular que muchas veces acompaña a esos ejercicios.
Fermentación láctica
En esta reacción el NADH se oxida y el ácido pirúvico se reduce transformándose en ácido láctico.
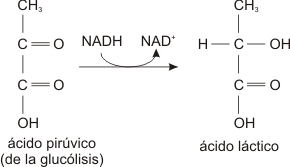
La fermentación sea ésta alcohólica o láctica ocurre en el citoplasma.
|
Esquema bioquímico del proceso de fermentación A) Alcohólica : 2 ácido pirúvico + 2 NADH Þ 2 etanol + 2 CO2 + 2 NAD+ B) Láctica : 2 ácido pirúvico + 2 NADH Þ 2 ácido láctico + 2 NAD+
|
La finalidad de la fermentación es regenerar el NAD+ permitiendo que la glucólisis continúe y produzca una provisión pequeña pero vital de ATP para el organismo.
RESPIRACIÓN AERÓBICA
En presencia de oxígeno, la etapa siguiente de la degradación de la glucosa es la respiración, es decir la oxidación escalonada del ácido pirúvico a dióxido de carbono y agua.
La respiración aeróbica se cumple en dos etapas: el ciclo de Krebs y el transporte de electrones y la fosforilación oxidativa (estos dos últimos procesos transcurren acopladamente).
En las células eucariotas estas reacciones tienen lugar dentro de las mitocondrias; en las procariotas se llevan acabo en estructuras respiratorias de la membrana plasmática.
Estructura de las Mitocondrias
Las mitocondrias están rodeadas por dos membranas, una externa que es lisa y una interna que se pliega hacia adentro formando crestas. Dentro del espacio interno de la mitocondria en torno a las crestas, existe una solución densa (matriz o estroma) que contiene enzimas, coenzimas, agua, fosfatos y otras moléculas que intervienen en la respiración.
La membrana externa es permeable para la mayoría de las moléculas pequeñas, pero la interna sólo permite el paso de ciertas moléculas como el ácido pirúvico y ATP y restringe el paso de otras. Esta permeabilidad selectiva de la membrana interna, tiene una importancia crítica porque capacita a las mitocondrias para destinar la energía de la respiración para la producción de ATP.
La mayoría de las enzimas del ciclo de Krebs se encuentran en la matriz mitocondrial. Las enzimas que actúan en el transporte de electrones se encuentran en las membranas de las crestas.
Las membranas internas de las crestas están formadas por un 80 % de proteínas y un 20 % de lípidos.
En las mitocondrias, el ácido pirúvico proveniente de la glucólisis, se oxida a dióxido de carbono y agua, completándose así la degradación de la glucosa.
El 95 % del ATP producido se genera, en la mitocondria.
Las mitocondrias son consideradas organoides semiautónomos, porque presentan los dos ácidos nucleicos (del tipo procarionte)
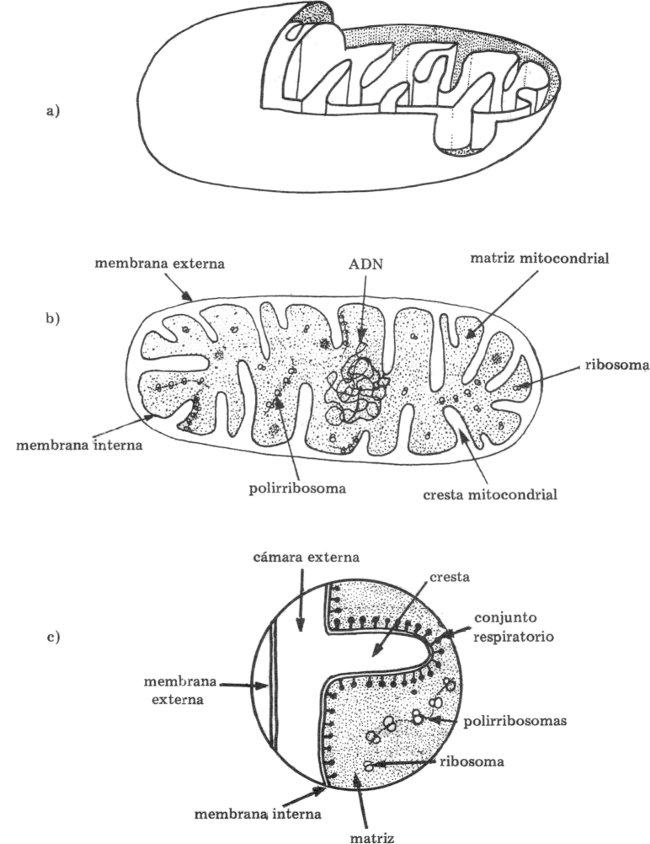
Fig. 9.2 - Esquema de la ultraestructura de una mitocondria. (a) Esquema tridimensional, (b) Esquema de un corte al M.E.T. (c) Cresta mitocondrial (detalle).
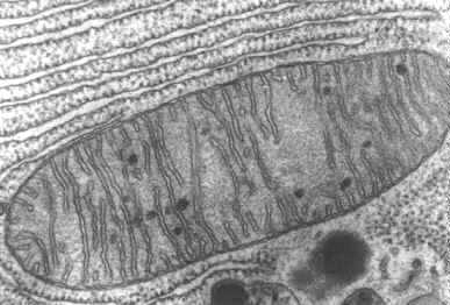
Fig.9.3- Microfotografía electrónica de una mitocondria. Se observan las invaginaciones de la membrana interna que forman las características crestas, que identifican esta organela
Como puede apreciase en la fig. 9.2 (c), las crestas mitocondriales aparecen cubiertas por partículas en forma de hongo, que tienen un tallo más fino que las unen a la membrana. Estas estructuras son las llamadas partículas F1 y representan una porción de la ATPasa especial que interviene en el acoplamiento entre la oxidación y la fosforilación. Las partículas F1 se encuentran en la membrana interna, del lado relacionado con la matriz; le confieren una asimetría característica relacionada con la función de la ATPasa (este punto se verá más detalladamente al referirnos a la hipótesis quimiosmótica).
Para concluir, es importante destacar que el ciclo de Krebs se lleva a cabo en la matriz mitocondrial; mientras que el transporte de electrones y la fosforilación oxidativa se producen a nivel de las crestas mitocondriales.
Ingreso al CICLO DE KREBS
El ácido pirúvico sale del citoplasma, donde se produce mediante glucólisis y atraviesa las membranas externa e interna de las mitocondrias. Antes de ingresar al Ciclo de Krebs, el ácido pirúvico, de 3 carbonos, se oxida. Los átomos de carbono y oxígeno del grupo carboxilo se eliminan como dióxido de carbono (descarboxilación oxidativa) y queda un grupo acetilo, de dos carbonos. En esta reacción exergónica, el hidrógeno del carboxilo reduce a una molécula de NAD+ a NADH.
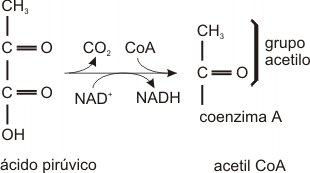
Ahora la molécula original de glucosa se ha oxidado a dos moléculas de CO2, y dos grupos acetilos y, además se formaron 4 moléculas de NADH (2 en la glucólisis y 2 en la oxidación del ácido pirúvico).
Cada grupo acetilo es aceptado por un compuesto llamado coenzima A dando un compuesto llamado acetilcoenzima A (acetil CoA). Esta reacción es el eslabón entre la glucólisis y el ciclo de Krebs.
CICLO DE KREBS
El ciclo de Krebs también conocido como ciclo del ácido cítrico es la vía común final de oxidación del ácido pirúvico, ácidos grasos y las cadenas de carbono de los aminoácidos.
La primera reacción del ciclo ocurre cuando la coenzima A transfiere su grupo acetilo (de 2 carbonos) al compuesto de 4 carbonos (ácido oxalacético) para producir un compuesto de 6 carbonos (ácido cítrico).
El ácido cítrico inicia una serie de pasos durante los cuales la molécula original se reordena y continúa oxidándose, en consecuencia se reducen otras moléculas: de NAD+ a NADH y de FAD+ a FADH2. Además ocurren dos carboxilaciones y como resultado de esta serie de reacciones vuelve a obtenerse una molécula inicial de 4 carbonos el ácido oxalacético.
El proceso completo puede describirse como un ciclo de oxalacético a oxalacético, donde dos átomos de carbono se adicionan como acetilo y dos átomos de carbono (pero no los mismos) se pierden como CO2.
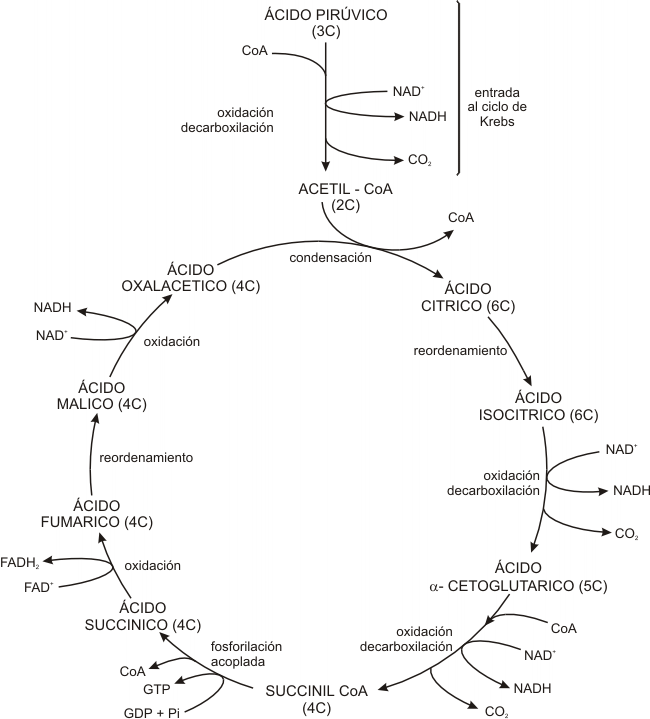
Fig. 9.4- Esquema simplificado del Ciclo de Krebs
Dado que por cada molécula de glucosa inicial se habían obtenido dos de ácido pirúvico y, por lo tanto dos de acetil CoA, deben cumplirse dos vueltas del ciclo de Krebs por cada molécula de glucosa. En consecuencia los productos obtenidos de este proceso son el doble del esquema que se detalla a continuación.
|
Cuadro 9.1 - Balance
parcial de la respiración
|
||
|
Proceso
|
Sustrato
|
Productos
|
|
Glucólisis
|
Glucosa
|
2 ácido pirúvico 2 ATP 2 NADH |
|
Entrada al ciclo de Krebs
|
2 ácido pirúvico
|
2 Acetil CoA 2 CO2 2 NADH |
|
Ciclo de Krebs
|
2 Acetil CoA
|
4 CO2 2 GTP (equivalentes a 2 ATP) 6 NADH 2 FADH2 |
|
Glucosa
|
|
|
Observando el balance parcial del ciclo de Krebs, se comprueba que en este proceso no se obtiene energía directamente bajo la forma de ATP (sólo se obtiene 1 GTP que es equivalente a 1 ATP). En cambio se obtienen cantidades de coenzimas reducidas (NADH y FADH2), y es a través de la oxidación posterior que se obtendrá la energía para sintetizar ATP.
Cada coenzima NADH equivale a 3 ATP y cada coenzima FADH2 equivale a 2 ATP.
TRANSPORTE DE ELECTRONES O CADENA RESPIRATORIA
En esta etapa se oxidan las coenzimas reducidas, el NADH se convierte en NAD+ y el FADH2 en FAD+. Al producirse esta reacción, los átomos de hidrógeno (o electrones equivalentes), son conducidos a través de la cadena respiratoria por un grupo de transportadores de electrones, llamados citocromos. Los citocromos experimentan sucesivas oxidaciones y reducciones (reacciones en las cuales los electrones son transferidos de un dador de electrones a un aceptor).
En consecuencia, en esta etapa final de la respiración, estos electrones de alto nivel energético descienden paso a paso hasta el bajo nivel energético del oxígeno (último aceptor de la cadena), formándose de esta manera agua.
Cabe aclarar que los tres primeros aceptores reciben el H+ y el electrón conjuntamente. En cambio, a partir del cuarto aceptor, sólo se transportan electrones, y los H+ quedan en solución.
FOSFORILACIÓN OXIDATIVA
El flujo de electrones está íntimamente acoplado al proceso de fosforilación, y no ocurre a menos que también pueda verificarse este último. Esto, en un sentido, impide el desperdicio ya que los electrones no fluyen a menos que exista la posibilidad de formación de fosfatos ricos en energía. Si el flujo de electrones no estuviera acoplado a la fosforilación, no habría formación de ATP y la energía de los electrones se degradaría en forma de calor.
Puesto que la fosforilación del ADP para formar ATP se encuentra acoplada a la oxidación de los componentes de la cadena de transporte de electrones, este proceso recibe el nombre de fosforilación oxidativa.
En tres transiciones de la cadena de transporte de electrones se producen caídas importantes en la cantidad de energía potencial que retienen los electrones, de modo que se libera una cantidad relativamente grande de energía libre en cada uno de estos tres pasos, formándose ATP.
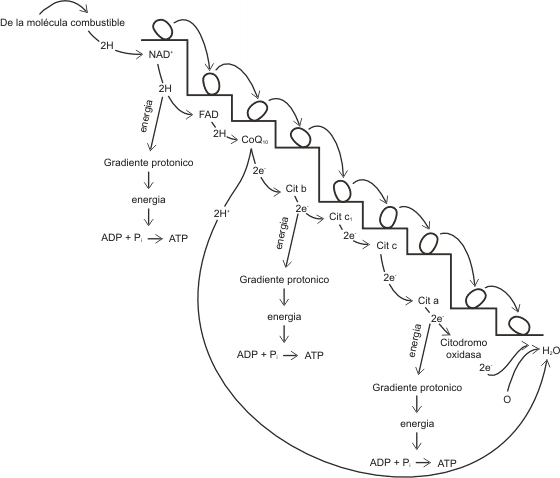
Fig.9.5- Diagrama de la cadena respiratoria y de la fosforilación oxidativa asociada
HIPÓTESIS QUIMIOSMÓTICA
Durante mucho tiempo se intentó explicar la naturaleza del enlace entre la cadena respiratoria y el sistema de fosforilación. En 1961, Mitchell propuso la hipótesis quimiosmótica, que es la que actualmente se acepta en general.
Esta hipótesis ha sido apoyada por las evidencias experimentales encontradas en distintos laboratorios, lo que le valió a Mitchell el premio Nobel en 1978.
La misma propone que el transporte de electrones y la síntesis de ATP están acopladas por un gradiente protónico a través de la membrana mitocondrial.
Según este modelo, el transporte de electrones paso a paso, desde el NADH o el FADH2 hasta el oxígeno a través de los transportadores de electrones, da por resultado el bombeo de protones a través de la membrana mitocondrial interna hacia el espacio entre las membranas mitocondriales interna y externa.
Este proceso genera un potencial de membrana a través de la membrana mitocondrial interna, ya que el medio que ocupa el espacio intermembranoso se carga positivamente.
La diferencia en concentración de protones entre la matriz y el espacio intermembranoso representa energía potencial, resultado en parte de la diferencia de pH y en parte de la diferencia en la carga eléctrica de los lados de la membrana. Cuando los protones pueden fluir de regreso a la matriz, descendiendo por el gradiente protónico, se libera energía utilizable en la síntesis de ATP a partir de ADP y Pi.
Los protones regresan a la matriz a través de conductos especiales situados en la membrana interna. Estos conductos están dados por un gran complejo enzimático, llamado ATP SINTETASA. Este complejo consta de dos proteínas: F0 y F1.
Las partículas F0 están incluidas en la membrana mitocondrial interna y la atraviesan desde afuera hacia adentro. Se presume que poseen un conducto o poro interior que permite el paso de los protones. Las partículas F1 (que ya habíamos mencionado, al describir la estructura mitocondrial) son proteínas globulares grandes consistentes en nueve subunidades polipeptídicas unidas a las partículas F0 en el lado de la membrana que linda con la matriz. Se comprobó que propulsa la síntesis de ATP a partir de ADP y Pi. Conforme los protones descienden a lo largo del gradiente de energía, dicha energía utiliza para sintetizar ATP. De esta manera, el gradiente protónico que existe a través de la membrana mitocondrial interna acopla la fosforilación con la oxidación.
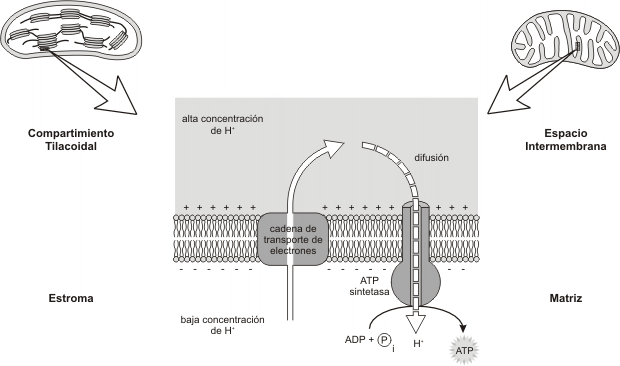
Fig. 9.6 - Esquema comparativo de la quimiósmosis en la mitocondria y el cloroplasto. Observe el bombeo de protones desde la matriz mitocondrial al espacio intermembrana (sombreado). El ATP se forma del lado de la membrana que mira a la matriz, por la difusión de los H+ a través del complejo ATPsintetasa. En el cloroplasto, a través de la membrana tilacoidal se bombean protones desde el estroma al compartimiento tilacoidal (sombreado). Como los H+ atraviesan la membrana a través de la ATPsintetasa, la fosforilación del ADP tiene lugar del lado de la membrana que mira al estroma.
|
Cuadro 9.2 - RESUMEN DE LA GLUCÓLISIS Y DE LA RESPIRACIÓN |
||
|
En el citoplasma: Glucólisis |
2 ATP |
2 ATP |
|
En las mitocondrias: De la glucólisis: De la respiración Ácido pirúvico Ciclo de Krebs: |
2 NADH 1
NADH 1 ATP 3 NADH 1 FADH2 |
|
Rendimiento total de ATP
|
||
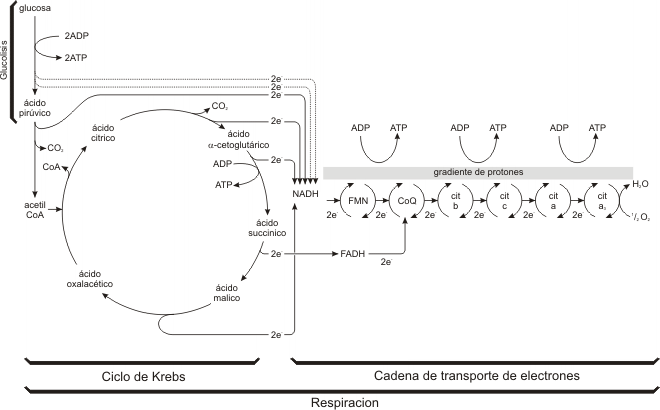
Fig. 9.7 - Resumen de la Glucólisis y de la Respiración. La glucosa se degrada a ácido pirúvico, en el citoplasma con un rendimiento de 2 moléculas de ATP y la reducción (flechas entrecortadas) de dos moléculas de NAD+ a NADH. El ácido pirúvico se oxida a acetil CoA y se reduce una molécula de NAD+, esta reacción y la siguiente ocurren 2 veces por cada molécula de glucosa (pasaje de e- con línea entera). En el ciclo de Krebs, el grupo acetilo se oxida y los aceptores de electrones NAD+ y FAD se reducen. El NADH y FADH2 transfieren sus electrones a la serie de transportadores de la cadena de transporte de electrones. Al circular los electrones hacia niveles energéticos menores se liberan cantidades relativamente grandes de energía libre . Esta liberación transporta protones a través de la membrana mitocondrial interna estableciendo el gradiente de protones que propulsa la síntesis de ATP a partir del ADP.
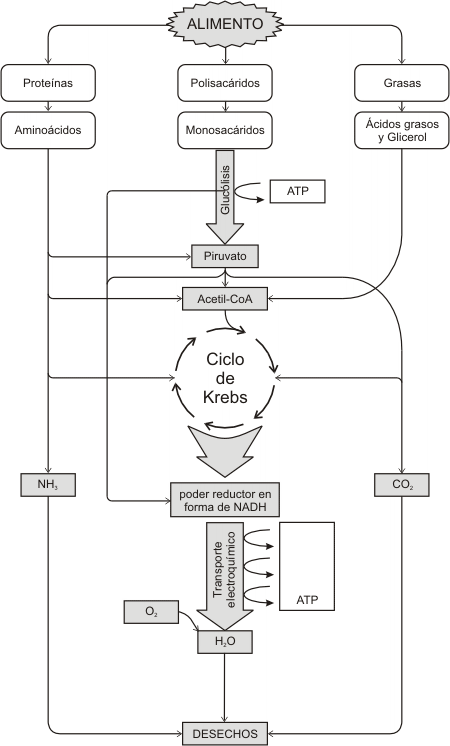
OTRAS VÍAS CATABÓLICAS
Sí la mayoría de los organismos no se alimentan directamente de glucosa. ¿cómo obtienen energía a partir de las grasas o proteínas?. La respuesta está en que el ciclo de Krebs es un gran nudo del metabolismo energético. Otras sustancias alimenticias son degradadas y convertidas en moléculas capaces de ingresar al ciclo.
Las grasas se desdoblan en sus componentes glicerol y ácidos grasos. Estos últimos son fraccionados en fragmentos de dos carbonos e introducidos en el ciclo de Krebs como acetil CoA.
Las proteínas se degradan a aminoácidos, estos son desaminados (se les eliminan los grupos amino) y el esqueleto de carbonos se convierte en un grupo acetilo, ingresando al ciclo de Krebs. Los grupos amino si no se utilizan, se excretan como urea u otros desechos nitrogenados
 .
.
Fig.9.8- Vías principales del catabolismo y anabolismo en la célula, Se observan las tres etapas, la primera tiene lugar en el lumen del tubo digestivo, la segunsa en el citosol y la última en las mitocondrias.
RESUMEN
En el afán de analizar detenidamente cada paso de las reacciones metabólicas de fotosíntesis y respiración, perdemos la noción de estos procesos globalmente.
En la fotosíntesis, la energía lumínica se convierte en química y se fija carbono en compuestos orgánicos.
Los fotosintetizadores o autótrofos elaboran hidratos de carbono a partir de CO2 y agua y liberan O2 a la atmósfera. Son estos organismos los que mantienen estables las concentraciones de CO2, y O2 atmosféricos.
En la respiración aeróbica los compuestos orgánicos son degradados a CO2 y H2O con la concomitante producción de energía química bajo la forma de ATP.
FOTOSÍNTESIS
En la primera etapa o etapa lumínica, la energía del sol es captada por la clorofila y otros pigmentos accesorios, provocando una serie de reacciones de óxido--reducción que propulsan la síntesis de ATP; la reducción de la coenzima NADP a NADPH y la oxidación de moléculas de H2O liberando O2 al medio. En la siguiente etapa o ciclo de Calvin el NADPH y el ATP (productos de la anterior etapa) se utilizan para reducir al CO2 que el vegeta1 toma del medio, a carbono orgánico. Si falta alguno de estos sustratos, el proceso se detiene.
Son necesarias 6 vueltas al c1clo para formar una molécula de glucosa partir de 2 moléculas de PGAL.
Este compuesto también se puede utilizar como material inicial para elaborar otros compuestos orgánicos que la célula necesita.
RESPIRACIÓN
La oxidación de la glucosa es una fuente principal de energía en la mayoría de las células.
La primera fase de este proceso es la glucólisis, en la cual la molécula de glucosa (6C), se escinde en dos moléculas de ácido pirúvico (3C). Este paso produce un rendimiento neto de 2 moléculas de ATP y dos moléculas de NADH.
La segunda fase de la degradación de la glucosa es la respiración aeróbica que ocurre en tres etapas: ciclo de Krebs, transporte de electrones y fosforilación oxidativa.
En ausencia deO2 el ácido pirúvico de la glucólisis se convierte en etanol o ácido láctico mediante fermentación. En el curso de la respiración las moléculas de ácido pirúvico se fraccionan en grupos acetilos; los cuales ingresan al ciclo de Krebs. En este ciclo los grupos acetilos se oxidan por completo a CO2, se reducen cuatro aceptores de electrones (tres NAD+ y Un FAD) y se forma GTP.
La etapa final de la respiración es el transporte de electrones y la fosforilación oxídativa (se dan acopladamente). En este paso intervienen una cadena de transportadores de electrones que transportan los electrones de alta energía aceptados por el NADH y el FADH2 viajando cuesta abajo hacia el oxígeno.
En tres puntos de su descenso por toda la cadena transportadora, se liberan grandes cantidades de energía que propulsan el bombeo de fotones hacía el espacio intermembranoso de la mitocondria. Esto crea un gradiente electroquímico a través de la membrana interna. Cuando los protones atraviesan el complejo ATP sintetasa hacia la matriz, la energía liberada se utiliza para sintetizar moléculas de ATP. Este mecanismo por el cual se cumple la fosforilación oxidativa se conoce como hipótesis quimiosmótica.
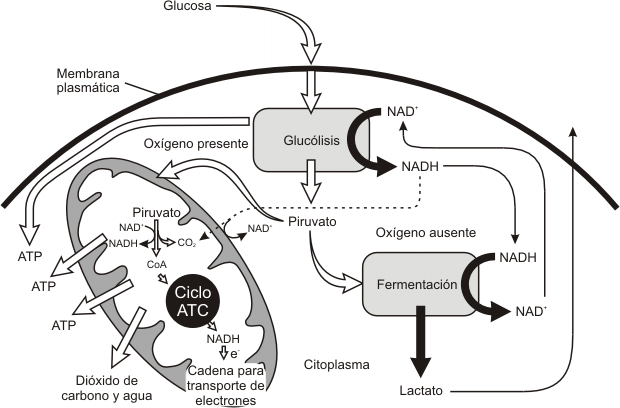
Fig.9.9- Resumen del metabolismo de los glúcidos en células eucariotas
ACTIVIDADES DE AUTOEVALUACIÓN
1) Esquematiza la estructura de una mitocondria y describe donde tienen lugar las diversas etapas de la degradación de la glucosa, en relación con estructura mitocondrial. ¿Qué moléculas e iones atraviesan las membranas mitocondriales en estos procesos?
2) Distingue lo siguiente: glucólisis / respiración / fermentación / vías aeróbias / vías anaerobias: FAD / FADH2; ciclo de Krebs / transporte de electrones.
3) Sigue una molécula de glucosa desde su ingreso a la célula hasta la formación de CO2 y H2O. Diferencia las etapas.
4) Complete el siguiente cuadro:
|
Proceso |
Glucólisis |
Respiración Aeróbica |
||
|
Ciclo de Krebs |
Cadena Respiratoria |
Fosforilación Oxidativa |
||
|
Ubicación |
||||
|
Sustrato |
||||
|
Producto |
||||
|
Ganancia |
||||
5) Al retirar de la membrana mitocondrial la porción F1 del complejo ATPsintetasa y estudiarla en solución, funciona como una ATPasa. ¿Por qué no funciona como una ATPsintetasa?
6) En caso de agotarse las reservas de glúcidos y lípidos. ¿A qué compuestos recurre la célula y a qué etapa del metabolismo se incorpora?.
6) Si la glucosa está constituida por: carbono, hidrógeno y oxígeno, explica:
a) ¿ Cuál es el destino del H+ que se desprende en el proceso?
b) ¿ En qué se transforman los átomos de C y O que se liberan?
c) ¿De dónde proviene la energía almacenada en la glucosa y liberada parcialmente en la glucólisis?
PREGUNTAS MULTIPLE OPCIÓN
1- En la siguiente reacción : " Piruvato + NADH + H+ Þ lactato + NAD":
a- el piruvato se reduce a lactato
b- el piruvato y NADH son reducidos a lactato y NAD
c- el piruvato se hidroliza a lactato
d- el NAD+ se reduce a NADH
e- el piruvato dona 2e- del lactato
2- La membrana externa de la mitocondria :
a- es más permeable que la interna
b- es menos permeable que la interna
c- es donde se localizan las proteínas de la cadena de transporte de electrones
d- sintetiza la matriz intermembranosa
e- presenta pliegues que proveen una mayor superficie de contacto
3- ¿Cuál de las siguientes reacciones es común a la respiración aeróbica y a la fermentación?:
a- malato Þ ácido oxalacético
b- fosfoenolpiruvato Þ piruvato
c- piruvato Þ lactato
d- piruvato Þ acetil CoA
e- fosfoenolpiruvato Þ ácido oxalacético
4- ¿Cuál de los siguientes compuestos no se encuentra en la matriz mitocondrial?
a- enzimas de la vía glucolítica
b- enzimas del ciclo de Krebs
c- ADN
d- Ribosomas
e- a y c son correctas
5- La b-oxidación de ácidos grasos :
a- es un proceso citosólico de síntesis
b- tiene menor rendimiento energético por mol de sustrato oxidado que la glucólisis aeróbica
c- se lleva a cabo en la matriz mitocondrial
d- es un proceso de síntesis peroxisomal
e- ninguna es correcta
BIBLIOGRAFÍA
l Alberts, B et al; (1996) Biología Molecular de la Célula. 3ra Edición. Ediciones Omega S.A. Barcelona.
l Campbell, N; (1997) Biology. 4th Edition. the Benjamin Cummings Publishing Company. Inc. California
l Castro, Handel y Rivolta . Actualizaciones en Biología. (1986). Ed. EUDEBA
l Castro R. et al. Investigación y Ciencia., N° 39, Diciembre de 1979.
l Curtis y Barnes (1992). Biología. 5ª Ed. Bs.As. Editorial Médica Panamericana.
l CONICET-SENOC (1984) Módulo 3 y 4.
l De Robertis(h); Hib; Ponzio. (1996).Biología Celular y Molecular de De Robertis. 12º Edición. El Ateneo. Bs.As.
l De Robertis, E.; Hib, J.; (1998) .Fundamentos de Biología Celular y Molecular. El Ateneo. Bs.As.
l Karp, G.; (1998) Biología Celular y Molecular. Ed. Mc Graw Hill Interamericana. México.
l Smith and Wood; (1997) .Biología Celular. Ed.Addison-Wesley, Iberoamericana S.A.
l Solomon y col. (1998) . Biología de Villee. 4ª. Ed. Mex. McGraw-Hill. Interamericana